Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ IV. МОЛЕКУЛЯРНІ МЕХАНІЗМИ СПАДКОВОСТІ ТА РЕАЛІЗАЦІЇ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ
ГЛАВА 20. МОЛЕКУЛЯРНІ МЕХАНІЗМИ РЕПЛІКАЦІЇ ДНК ТА ТРАНСКРИПЦІЇ РНК
20.3. МОЛЕКУЛЯРНІ МЕХАНІЗМИ РЕПЛІКАЦІЇ ДНК
Загальна схема процесу біосинтезу ДНК, що була вивчена в системі А. Корнберга, залишилася правильною до даного часу. Але, враховуючи, що молекула ДНК є подвійною спіраллю із закрученими один навколо одного антипаралельними ланцюгами, існують певні геометричні — топологічні складності, що виникають при реплікації молекул ДНК прокаріотів та еукаріотів.
Топологічні проблеми реплікації ДНК
1. Спіралізація та суперспіралізація подвійної спіралі ДНК. Топоізомерази.
Нативні ДНК є двоспіральними молекулами, тому реплікації материнської молекули повинно передувати її розплетення з утворенням реплікативної вилки, що складається з двох розведених матричних ланцюгів (відповідно до напівконсервативного механізму реплікації). Структурні складності цього процесу полягають також у тому, що у прокаріотів (зокрема у E. Coli) двоспіральні молекули ДНК є кільцевими, зачіпленими — тобто вони не мають вільних кінців, і їх неможливо розділити без розриву одного з ланцюгів.
Розплетення та відокремлення двох ланцюгів ДНК, що передує синтезу дочірніх ланцюгів, реалізується при дії спеціальних типів білків:
- ферментів топоізомераз, які забезпечують зміну кількості супервитків у кільцевих замкнутих молекулах ДНК. Топоізомерази, і, зокрема ДНК-гіраза прокаріотів (gyration — обертання, англ.), здійснюють у зачіплених нитках ДНК розриви полінуклеотидних ланцюгів, що створює механічні передумови для їх розкручування. Топоізомерази присутні також у хроматиновому апараті еукаріотів: вони вносять дволанцюгові розриви в довгі лінійні ДНК хромосом, забезпечуючи підготовку останніх до реплікації;
- ферментів хеліказ(helix — спіраль, англ.), що в АТФ-залежному процесі розкручують, розплітають короткі ділянки ДНК, утворюючи реплікативні вилки(replication fork — англ.) — місця послідовного розплетення подвійного ланцюга та синтезу нових ниток ДНК;
- білків, що зв’язують однониткові ДНК(SSB-білки — single strand binding proteins, англ.), протидіючи їх повторному об’єднанню (ренатурації).
Утворення реплікативної вилки в кільцевих дволанцюгових молекулах ДНК E. Coli призводить до формування тета-структур (таких, що нагадують грецьку літеру «тета»), які містять реплікативну вилку, що пересувається у міру синтезу дочірніх ланцюгів ДНК:

Рис. 20.7. Кільцева хромосома E. Coli в період реплікації ДНК. Геометрична структура, що утворилася, нагадує грецьку літеру θ («тета»)
(за L. Stryer, 1995; модифіковано).
У ДНК еукаріотів, що організовані у вигляді лінійних дволанцюгових молекул у складі ядерного хроматину, утворюється водночас багато (ймовірно, від сотень до декількох тисяч) реплікативних вилок, що сприяє ефективній реплікації цілої еукаріотичної хромосоми. Одночасна реплікація ДНК в багатьох точках морфологічно виявляється утворенням упродовж хромосоми «реплікаційних бульбашок» (рис. 20.8).
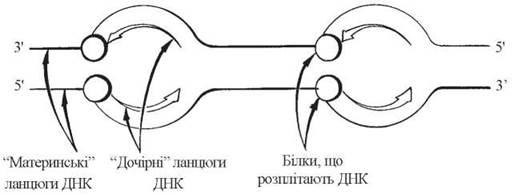
Рис. 20.8. Утворення реплікаційних «бульбашок» у процесі подвоєння хромосом еукаріотів (за D. Granner, 1988; модифіковано).
Ділянки геному еукаріот, де формуються реплікативні вилки, мають назву «точок ori» (origin — початок, англ.). Завдяки багатоцентровій реплікації хромосоми, повне подвоєння генетичного матеріалу клітин вищих організмів (зокрема, ссавців) займає близько 9 год.
2. Значення антипаралельності ланцюгів ДНК. Фрагменти Оказакі
У зв’язку з антипаралельністю двох ланцюгів ДНК (один — 5'→3', а другий — 3'→5'), одночасна реплікація комплементарних їм ланцюгів повинна була б йти також у протилежних напрямках(тобто 3'→5' та 5'→3', відповідно). Але, як уже наголошувалося, ДНК-полімерази можуть синтезувати полідезоксирибонуклеотидні ланцюги лише в напрямку 5'→3'. Це протиріччя було усунуте завдяки дослідженням японського вченого Реджі Оказакі (Okazaki), який встановив, що синтез одного з дочірніх ланцюгів ДНК є перервним і здійснюється у вигляді коротких (1-2 тис. нуклеотидів) ланцюгів, які пізніше з’єднуються між собою.

Рис. 20.9. Оказакі (Okazaki) Реджі (1930-1975).
Таким чином, синтез двох дочірніх ланцюгів ДНК здійснюється за різними механізмами. Розрізняють (рис. 20.10):
лідируючий (leading — англ.) ланцюг — який утворюється шляхом безперервного нарощування нуклеотидного ланцюга в напрямку 5'→3';
відстаючий (lagging — англ.) ланцюг — який утворюється з фрагментів Оказакі, що синтезуються в ДНК-полімеразних реакціях в напрямку 5'→3'.
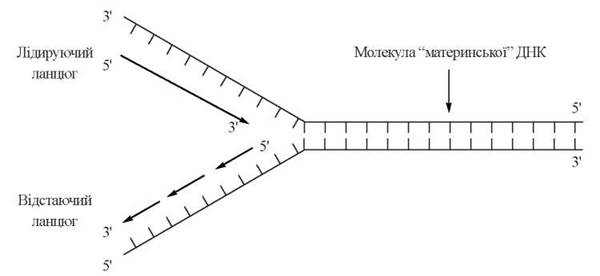
Рис. 20.10. Схема подвоєння ланцюгів ДНК за Р. Оказакі.
Етапи синтезу дочірніх ланцюгів молекул ДНК
1. Ініціація синтезу полідезоксирибонуклеотидних ланцюгів ДНК, якій передує утворення «затравних» (праймерних) ланцюгів РНК (РНК-праймерів), до 3'-ОН-груп яких здатні приєднуватися дНМФ, що утворюють нові (дочірні) ланцюги ДНК. Довжина цих праймерних ланцюгів складає в середньому від 10 до 200 нуклеотидів. Синтез РНК-праймерів відбувається за участю ферментів РНК-полімераз — так званих праймаз.
2. Елонгація синтезу ДНК, яка відбувається за різними механізмами на лідируючому та відстаючому ланцюгах.
2.1. На лідируючому ланцюгу нарощування дНМФ здійснюється ДНК-полімеразою III, що функціонує безперервно, утворюючи ланцюг ДНК від РНК-праймера до реплікативної вилки.
2.2. На відстаючому ланцюгу:
а) спочатку при дії ДНК-полімерази III синтезуються окремі фрагменти Оказакі, кожен з яких починається з відповідного РНК-праймера і закінчується перед початком передуючого йому РНК-праймера;
б) після формування фрагментів Оказакі ДНК-полімераза I, за рахунок своєї 5'→3'-екзонуклеазної активності видаляє РНК-праймери і (за рахунок 5'→3'-полімеразної активності) заміщує їх фрагментами ДНК;
в) розриви між окремими фрагментами Оказакі зшиваються спеціальним ферментом ДНК-лігазою.
Зазначені етапи реплікації ДНК подані на рис. 20.11.
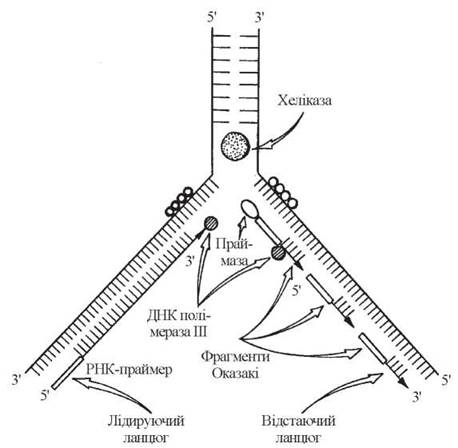
Рис. 20.11. Участь ферментів в реплікації ДНК шляхом синтезу лідируючого та відстаючого ланцюгів.
Таким чином, в результаті дії комплексу реплікативних білків, зчитування інформації з двох материнських ланцюгів ДНК з антипаралельною спрямованістю (5'→3' та 3'→5') супроводжується утворенням двох дочірніх ланцюгів з відповідними антипаралельними напрямками фосфодіефірних зв’язків (3'→5' та 5'→3', відповідно). Взаємодія комплементарних одного материнського та одного дочірнього ланцюгів призводить до формування двох молекул ДНК, як це передбачено напівконсервативним механізмом реплікації.
Розглянута схема реплікації ДНК, що була первинно досліджена на прокаріотах, є справедливою також і для еукаріотичних клітин. У цьому випадку також відбувається диференційований та протилежно спрямований синтез двох дочірніх ланцюгів — лідируючого та відстаючого. Безперервний синтез полідезоксирибонуклеотиду на лідируючому ланцюгу каталізується ДНК-полімеразою δ, а перервний на відстаючому ланцюгу — полімеразою α. Праймази утворюють тимчасові «затравні» РНК-ланцюги, які пізніше видаляються.