Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ IV. МОЛЕКУЛЯРНІ МЕХАНІЗМИ СПАДКОВОСТІ ТА РЕАЛІЗАЦІЇ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ
ГЛАВА 22. РЕГУЛЯЦІЯ ЕКСПРЕСІЇ ГЕНІВ. ГЕНЕТИЧНІ РЕКОМБІНАЦІЇ
22.2. ОСОБЛИВОСТІ МОЛЕКУЛЯРНОЇ ОРГАНІЗАЦІЇ ТА ЕКСПРЕСІЇ ГЕНОМУ В ЕУКАРІОТІВ
Як свідчать дані сучасної біохімії і молекулярної біології, склад та субклітинна архітектура еукаріотичної клітини суттєво відрізняються від прокаріотів: так, кількість індивідуальних білків в організмі людини перевищує 50 000, порівняно з приблизно 3 000 різних білків у E.Coli. Відповідно, і геном ядерних організмів є більш складною системою за своєю структурою та молекулярною організацією, порівняно з прокаріотами, зокрема значна різниця існує в кількісних показниках і структурній організації ДНК.
Відповідно до особливостей структурної організації геному, контроль експресії генів у еукаріотів також значно складніший. Окрім механізмів, близьких до існуючих у прокаріотів, в контролі генної експресії в еукаріотичних клітинах беруть участь такі специфічні молекулярні процеси, що реалізують регуляторну дію на різних рівнях генної експресії:
(1) на рівні структурної організації геному — регуляція здійснюється за рахунок наявності специфічних послідовностей нуклеотидів, генних перебудов (рекомбінації генів), ампліфікації генів;
(2) на рівні транскрипції — регуляторними механізмами є вплив сигналів посилення та послаблення транскрипції (енхансерів та атенюаторів, відповідно), постранскрипційна модифікація мРНК;
(3) на рівні трансляції — основним механізмом регуляції є ковалентна модифікація білкових факторів трансляції шляхом їх зворотного фосфорилювання — дефосфорилювання.
Молекулярна організація ДНК еукаріотів
Гаплоїдний геном кожної клітини організму Homo sapiens містить 3,5 · 109 пар азотистих основ і складається з 23 пар хромосом (рис. 22.7), що є достатнім для утворення близько 1,5 млн. пар генів. Разом з тим, реально в організмі людини синтезується не більше 100 000 різних білків, тобто більша частина ядерної ДНК, що складає геном людини, не транслюється в амінокислотну послідовність білкових молекул.

Рис. 22.7. Будова 12-ї хромосоми каріотипу людини (х 27850).
Наявність (крім регуляторних і сигнальних нуклеотидних послідовностей, присутніх також у прокаріотів) значної кількості ділянок, що не транскрибуються, є специфічною особливістю структури геному еукаріотичних клітин. Такі «мовчазні» фрагменти геному отримали назву інтронів, на відміну від екзонів — ділянок геному, які транскрибуються з утворенням мРНК, що несуть інформацію для синтезу специфічних клітинних білків.
Згідно з сучасними оцінками, тільки 2 % ДНК клітин ссавців містять інформацію для кодування білків організму (L. Stryer, 1995).
Послідовності ДНК, що повторюються
Вивчення нуклеотидних послідовностей ДНК вищих організмів виявило, крім унікальних за нуклеотидним складом послідовностей, розповсюдженість фрагментів, що присутні, зокрема в ДНК ссавців, у вигляді багатьох копій — від 2 до 107 повторів на клітину. В геномі людини повтори складають 20-30 % від його загальної довжини.
Залежно від кількості нуклеотидів і кратності їх повторення послідовності нуклеотидів, що повторюються, підрозділяються на такі класи:
(а) високоповторні послідовності, які є ділянками з довжиною від 5 до 500 пар нуклеотидів, розташованими один за одним («тандемно»). Такі послідовності утворюють кластери, що містять від 1 до 10 млн. копій і складають фракцію «сагелттної ДНК». Ці високоповторні послідовності транскрипційно неактивні, і їх функціональну роль остаточно не з’ясовано;
(б) помірноповторні послідовності, які мають не більше ніж 106 копій і не утворюють тандемних кластерів, а чергуються з унікальними нуклеотидними послідовностями («дисперговані повтори»). Ці послідовності мають різну довжину і можуть бути як короткими, так і довгими (в 5-7 тис. пар нуклеотидів).
Короткі дисперговані повтори — це фрагменти ДНК довжиною від одиниць (декількох пар) до декількох сотень пар нуклеотидів. До цього класу належать поширені в клітинах організму людини повтори сімейства Alu, які мають довжину 300 пар нуклеотидів і повторюються в кількості близько 500 000 копій, складаючи в цілому 3-6 % від загального розміру геному гаплоїдної клітини. Ці повтори можуть транскрибуватися у вигляді компонентів гяРНК та індивідуальних клітинних РНК — 4,5s- і 7s-РНК.
За своєю нуклеотидною структурою повтори Alu близькі до кінцевих послідовностей ретровірусів LTR і, ймовірно, мають з ними генетичний зв’язок. Вважають, що ці послідовності є мобільними елементами геному, що здатні до транспозиції, тобто вирізування і вбудови в різні ділянки геному.
Ядерний хроматин еукаріотів
Як уже зазначалося, структурною особливістю просторової будови ДНК еукаріотів є її компактизація у вигляді суперспіралей, які, в комплексі з гістонами та негістоновими білками — НГБ (до їх складу входять регуляторні, структурні білки, ферменти реплікації тощо) утворюють ядерний хроматин, що на етапі мітозу організується у вигляді хроматинових тілець — хромосом.
У клітинах тварин та людини реплікація ДНК відбувається тільки в певний період клітинного циклу — S-фазу, яка відокремлена від мітозу (М) пресинтетичною (G1) та постсинтетичною (G2) фазами; фази S, G1 та G2 складають «інтермітотичний період» — інтерфазу, G0 — період репродуктивного спокою (рис. 22.8). Контрольні механізми, що регулюють перехід клітини з G1 до S-фази залишаються недостатньо з’ясованими. Разом з тим, пускові реакції, які визначають початок мітозу, і полягають в зворотному фосфорилюванні певних ядерних та цитоплазматичних білків, протягом останніх років в достатній мірі розшифровані (див. нижче).
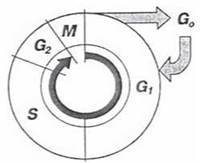
Рис. 22.8. Фази клітинного циклу.
Нуклеосомна організація ядерного хроматину та біохімічні характеристики гістонів були розглянуті в главі 3. В інтерфазному ядрі хроматин диференціюється на транскрипційно активний хроматин (ТАХ), або «еухроматин» та транскрипційно неактивний, репресований хроматин (РХ), або «гетерохроматин». Ці два типи хроматину розрізняють за біохімічними та морфологічними (електронно-мікроскопічними) характеристиками: ТАХ менш щільно компактизований, ніж РХ, його нуклеосомна структура змінена, а в транскрипційно найбільш активних регіонах взагалі відсутня.
Ковалентна модифікація гістонів та НГБ
Гістони та НГБ, що входять до складу нуклеосом хроматину, підлягають процесам посттрансляційної ковалентної модифікації, яка змінює їх хімічні властивості, зокрема здатність до взаємодії з певними ділянками ДНК, і може бути одним з біохімічних механізмів контролю ступеня експресії певних генів.
До реакцій ковалентної модифікації білків ядерного хроматину належать їх ацетилювання, фосфорилювання, метилювання, глікозилювання, АДФ-рибозилювання. Негістонові білки підлягають постсинтетичним модифікаціям в значно більшому ступені, ніж гістони. Відповідні радикали (ацетильні, фосфорильні, метильні, глікозильні, АДФ-рибозильні) сполучаються з боковими ланцюгами амінокислотних залишків в поліпептидних ланцюгах гістонів та НГБ.
Слід визнати, що регуляторне значення в реалізації конкретних молекулярно-генетичних процесів точно не встановлене ні для одного із зазначених шляхів ковалентної модифікації ядерних білків. Певні функціональні кореляції з активністю геному виявлені лише для реакцій ацетилювання та фосфорилювання.
Ацетилювання гістонів - процес, що починається вже в цитоплазмі (ацетилювання N-кінцевого серину гістону Н4) і завершується в ядрі, де ацетилюються переважно бокові радикали лізинових залишків, що призводить до зменшення позитивного заряду гістонів і розрихлення комплексів гістон-ДНК у складі нуклеосом. Ацетилювання передує активації геному і зростанню швидкості транскрипції.
Фосфорилювання - дефосфорилювання білків хроматину (гідроксильних груп залишків серину та треоніну) — клас реакцій, що відбуваються як у цитоплазмі (безпосередньо після їх трансляції), так і в ядрі. Встановлено, що протягом різних фаз клітинного циклу зворотно фосфорилюються як гістони (Н1, Н3), так і окремі НГБ, проте для індивідуальних білків ці закономірності можуть бути різними.
Генетичні рекомбінації
Важливе місце в організації геному різних клітин посідають генні перебудови (реаранжування — rearrangements, англ.), або генетичні рекомбінації (рекомбінації генів), під якими розуміють обмін фрагментами ДНК між різними генами або об’єднання генів з різних біологічних джерел з утворенням нових хромосомних структур, здатних до реплікації і генетичної експресії (транскрипції та трансляції).
Генетичні рекомбінації мають місце в біологічних системах різного ступеня складності — від вірусів і бактеріофагів до вищих еукаріотичних організмів. Перебудови структури генів є рушійною силою мінливості і мають велике значення як у формуванні біохімічної індивідуальності на рівні окремих особин в межах біологічного виду, так і в утворенні видів у процесі еволюції.
Молекулярні механізми генетичних рекомбінацій складні і суттєво відрізняються у різних організмів та при різних типах рекомбінацій. Вони включають у себе ферментативні процеси «розрізання» молекул реципієнтних ДНК (за участю специфічних ДНК-аз) та включення в молекулу ДНК чужорідних полінуклеотидних фрагментів з іншої хромосоми, або з іншого локусу тієї ж хромосоми — так званих транспозонів (transpose — переміщати, англ.). Здатність транспозонів бути вбудованими в молекули інших ДНК залежить від присутності на кінцях транспозонів особливих нуклеотидних фрагментів — так званих інсерційних послідовностей (insert — вставляти, включати, англ.); наступне зшивання фрагментів ДНК-лігазами завершує процес перенесення гена (рис. 22.9).
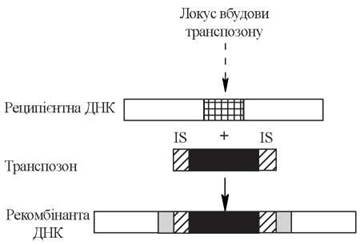
Рис. 22.9. Вбудова транспозону в рецитєнтну ДНК. На обох кінцях транспозону розміщені інсерційні послідовності (IS-елементи).
Рекомбінації у прокаріотів
Рекомбінації є поширеним процесом у прокаріотичних організмів, завдяки якому клітини останніх включають до складу свого геному фрагменти ДНК (генетичні елементи) з інших клітин. До генетичних рекомбінацій у прокаріотів належать:
а) трансформація — процес включення в геном реципієнтного мікроорганізму ДНК з донорної загиблої клітини того ж виду;
б) трансдукція — перенос бактеріофагом фрагмента ДНК інфікованої клітини в склад геному іншого реципієнтного мікроорганізму;
в) кон’югація — процес статевого розмноження, що має місце у деяких видів бактерій і полягає в перенесенні фрагмента ДНК з донорної (F+) до реципієнтної (F-) клітини.
Рекомбінації в еукаріотів
У вищих організмів генетичні рекомбінації є важливим елементом утворення гаплоїдних яйцеклітин та сперматозоїдів в процесі мейозу. Перед вступом клітин до редукційного поділу гомологічні хромосоми обмінюються ділянками ДНК (кросинговер), що є молекулярно-генетичним механізмом надбання нащадками властивостей обох батьківських особин.
Рекомбінації генів імуноглобулінів
У зрілих особин процеси переміщення (транслокації, транспозиції) окремих генів або груп генів в інше місце геному (в межах тієї ж хромосоми або в іншу хромосому) мають місце в генах В-лімфоцитів, що кодують утворення імуноглобулінів. Ці генетичні рекомбінації є механізмом, завдяки якому в організмі людини та тварин забезпечується синтез мільйонів різних антитіл у відповідь на проникнення в організм чужорідних антигенів (глава 30).
Імуноглобуліни є білками тетрамерної будови, що складаються з чотирьох полі- пептидних ланцюгів: двох H- («важких») ланцюгів та двох L- («легких») ланцюгів, які є попарно однаковими. Розрізняють п’ять типів H-ланцюгів (α — альфа, γ — гамма, μ — мю, δ — дельта, ε — епсилон) та два типи L-ланцюгів (k — каппа та λ —лямбда). Залежно від типу легкого та важкого ланцюгів, розрізняють п’ять класів імуноглобулінів (IgG, IgA, IgM, IgD та IgE), у складі кожного з яких певний тип H-ланцюга сполучається з одним із двох типів L-ланцюга.
У свою чергу, в кожному з H- та L-ланцюгів молекул імуноглобулінів можна виділити окремі структурні домени: константні (С-) ділянки, що розміщуються з С-кінців H- та L-ланцюгів і характеризуються сталістю амінокислотного складу в різних класів імуноглобулінів та варіабельні (V-) ділянки, що розміщуються з N-кінців H- та L-ланцюгів, і характеризуються мінливістю амінокислотного складу, утворюючи конформації, комплементарні детермінантним групам антигенів, тобто забезпечуючи їх зв’язування з антитілами.
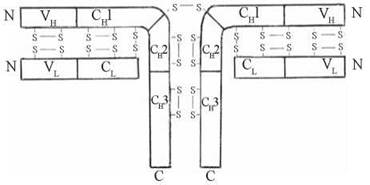
Рис. 22.10. Схема будови молекули імуноглобуліну.
Саме завдяки наявності на кінцях L- та H-ланцюгів імуноглобулінів варіабельних ділянок забезпечується можливість синтезу надзвичайно великої кількості індивідуальних імуноглобулінів (антитіл) відповідно до надходження чужорідних білкових молекул. У свою чергу, синтез такої значної кількості молекул з різною первинною структурою визначається множинними рекомбінаціями генів, що кодують окремі частини молекул імуноглобулінів.
Розглянемо основні закономірності генетичних рекомбінацій, що існують при формуванні в зрілих лімфоцитах генів, які кодують синтез L- та H-ланцюгів імуноглобулінів.
L-ланцюги — синтез легких ланцюгів відбувається за рахунок експресії трьох сімейств генів, що утворюють варіабельний (VL), з’єднувальний (JL) та константний (CL) сегменти; до того ж, зазначені три сімейства, що кодують синтез легких ланцюгів типу каппа, локалізовані в 2-й хромосомі, ланцюгів типу лямбда — в 22-й хромосомі.
У гаплоїдному хромосомному наборі міститься близько 500 генів сімейства VL, 56 генів типу JL та 10-20 генів типу CL. Ці генні сімейства локалізовані на відстані одне від одного в різних ділянках хромосом (2-ї та 22-ї, відповідно) і зближуються між собою в період утворення зрілого В-лімфоцита за рахунок перебудови (rearrangement) ДНК і транслокації гена типу VL з дистальної ділянки хромосоми ближче до JL- та CL-сегментів. За рахунок такої перебудови утворюється єдиний генний локус, що має будову VL(i)JL(i)CL(i) і транскрибується у вигляді єдиної поліцистронної мРНК-попередника (первинного транскрипту), який після відповідного процесингу утворює зрілу мРНК для легкого ланцюга імуноглобулінів.

Рис. 22.11. Генні сімейства, що утворюють Н-ланцюги імуноглобулінів (Stryer L., 1995).
H-ланцюги — синтез важких ланцюгів імуноглобулінів кодується генами чотирьох сімейств, що утворюють VH-, D-, JH- та CH-сегменти у складі 14-ї хромосоми:
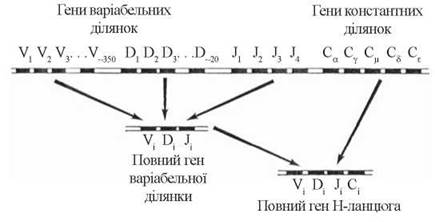
Генетичний локус, що кодує варіабельну ділянку важкого ланцюга, формується за рахунок зближення генів із сімейств VH (250-350 генів), D (15-20 генів) та JH (4 гени). Генна детермінація синтезу цілісного важкого ланцюга відбувається за рахунок взаємодії локусу будови VH(i)D(i)JH(i) (що кодує варіабільну ділянку) з одним з восьми генів сімейства СH (що кодують константну ділянку).
Синтез лімфоцитами імуноглобулінів певних класів (G, A, M, D, E), що розрізняються за типом будови важкого ланцюга, забезпечується за рахунок функціонування механізму «переключення класів важких імуноглобулінів», який здійснюється шляхом фіксації в структурі геному певного з обраних VH-генів з певним СH-геном. Такі генні перебудови і лежать в основі синтезу важких ланцюгів імуноглобулінів під час диференціювання ембріональних лімфоцитів (А.Я. Николаев, 1998).
Таким чином, завдяки можливостям утворення різних сполучень трьох генів сімейств VL, JL та CL при синтезі легких ланцюгів та чотирьох генів сімейств VH, D, JH, та СH при синтезі важких ланцюгів, в імунній системі формуються клони лімфоцитів з різноманітною бібліотекою генів, які забезпечують експресію величезної кількості імуноглобулінів (до 107-108) з різною антигенною специфічністю та функціональними властивостями.
Ампліфікація генів
Ампліфікація генів — процес збільшення кількості копій відповідних генів.
Молекулярною основою ампліфікації є багаторазова («вибухоподібна») ініціація синтезу ДНК (реплікації) в тому ж самому реплікаційному сайті (рис. 22.12).
Рис. 22.12. Схема процесу ампліфікації генів.
Прикладами ампліфікації генів вищих організмів є:
(1)ампліфікація генів металотіонеїну — білка, що зв’язує токсичні для організмів ссавців іони важких металів, зокрема ртуті, міді, цинку та кадмію. При надходженні в організм зазначених іонів відбувається збільшення частоти транскрипції металотіонеїнового гена в десятки разів, що є фізіологічним механізмом детоксикації важких металів;
(2) ампліфікація гена дигідрофолатредуктази — ферменту, що перетворює фолієву кислоту до її коферментних форм — дигідрофолатів, які є коферментами в синтезі пуринів та тиміну.
Ампліфікація гена дигідрофолатредуктази і збільшення в сотні разів рівня синтезу ферменту спостерігаються в умовах введення в організм онкологічних хворих препарату метотрексату. В основі антипухлинної дії метотрексату лежить інгібірування активності дигідрофолатредуктази, що призводить до порушення синтезу нуклеїнових кислот у клітинах злоякісних пухлин. Результатом такого активованого синтезу ферменту внаслідок ампліфікації гена є втрата чутливості клітин-мішеней до протипухлинного лікарського засобу.
Ланцюгова полімеразна реакція
Явище ампліфікації генів набуло важливого практичного застосування в методі ланцюгової полімеразної реакції (ЛПР), що був запропонований в 1983 р. американським дослідником К. Мюллісом (K. Mullis), і дістав поширення в сучасних біомедичних дослідженнях.
Метод ЛПР дозволяє отримати в чистому вигляді і в значній кількості (достатній для проведення досліджень) певні фрагменти (послідовності) ДНК складного геному людини та ідентифікувати їх за нуклеотидним складом. Принцип методу полягає в застосуванні РНК-праймерів, специфічних до певних ділянок ДНК, що аналізується, та подальшому використанні ДНК-полімерази, яка здатна реплікувати нуклеотидні послідовності в обох комплементарних ланцюгах, утворюючи значну кількість копій саме досліджуваного полінуклеотидного фрагмента. Ампліфіковані фрагменти ДНК виділяють з реакційного середовища і досліджують за допомогою існуючих біохімічних методів.
За допомогою ЛПР здійснюється так звана «ДНК-діагностика», тобто аналіз певних послідовностей ДНК в клітинах людини, що має першорядне значення в діагностиці спадкових хвороб, виявленні присутності в організмі людини певних вірусів (в тому числі ВІЛ), ідентифікації особистості. Так, зокрема, саме за цим методом був здійснений судово-медичний аналіз залишків царської сім’ї та ідентифіковані залишки тіла російського царя Миколи II.
Регуляція експресії генів еукаріотів на рівні транскрипції
Гени вищих організмів мають розвинену систему сигналів регуляції транскрипції, які не лише вказують на місце початку синтезу РНК, а й регулюють його активність. До складу системи транскрипційних сигналів у еукаріотів входять:
(1) промотори, специфічні для різних РНК-полімераз, але такі, що мають спільну нуклеотидну послідовність (TATA...), яка гомологічна блоку Прибнова у прокаріотів; з цим нуклеотидним блоком взаємодіє РНК-полімераза II;
(2) специфічні послідовності матричної ДНК, які підсилюють або послаблюють рівень експресії структурних генів, впливаючи на активність транскрипції, тобто на кількість молекул мРНК, що синтезуються, та швидкість їх утворення — «енхансерні» (підсилювальні), «аттенюаторні» (послаблювальні), «сайленсерні» (заглушувальні), «адапторні» елементи геному.
Енхансери
Найбільш вивченими на даний час є активуючі елементи системи регуляції транскрипції — позитивні регулятори енхансери (enhancers — посилювачі, англ.). Енхансери — це ділянки ДНК, які можуть складатися з нуклеотидних послідовностей (довжиною в декілька десятків пар азотистих основ — «модулей», або "мотивів» — motifs, англ.), що тандемно повторюються.
Енхансери збільшують ефективність транскрипції генів, на які вони впливають, в десятки і сотні разів. Встановлені нуклеотидні послідовності енхансерів для багатьох ферментних білків (хімотрипсину, алкогольдегідрогенази тощо), гормонів (інсуліну, плацентарного лактогену людини), імуноглобулінів.
Енхансери здатні впливати на активність відповідних генів навіть за умов віддаленості від їх промоторів на декілька тисяч азотистих основ. Активуюча дія енхансерів опосередкована регуляторними білками (див. нижче), що взаємодіють з ними і можуть впливати на траскрипцію віддалених ділянок ДНК. Разом з тим, при взаємодії з певними білковими факторами деякі енхансери можуть отримувати якості негативних регуляторів — сайленсерів (заглушувачів — англ.), що гальмують експресію відповідних генів.
(3) численні регуляторні білки, що є компонентами системи транскрипційних сигналів, контролюючи активність синтезу мРНК різних класів. На даний час виділені білки, що здатні до специфічної дії з певними енхансерами. Ці білки можуть взаємодіяти з регуляторними послідовностями ДНК, що розташовані на відстані в декілька тисяч азотистих основ від сайтів взаємодії з РНК-полімеразою та ініціації транскрипції. Вважають, що такий дистантний вплив регуляторних білків реалізується за рахунок змін у просторовій конформації ланцюгів ДНК і утворення внутрішніх петель, що зближають регуляторні елементи геному з ділянками, які транскрибуються.
В адаптації організму вищих тварин до умов середовища, що змінюються, важливу участь беруть стероїдні гормони кори наднирникових залоз. Встановлено, що дія цих фізіологічно активних сполук опосередкована специфічними білками — рецепторами, які в комплексі з гормонами взаємодіють з певними енхансерними ділянками геному, активуючи транскрипцію ДНК з певних генів.
Регуляція експресії генів еукаріотів на рівні трансляції
Механізмом регуляції біосинтезу білка на рівні трансляції є ковалентна модифікація білкових факторів трансляції шляхом зворотного фосфорилювання — дефосфорилювання за участю цАМФ-залежного регуляторного каскаду клітини.
Контроль трансляції шляхом фосфорилювання (інактивації) та дефосфорилювання (активації) фактора ініціації трансляції eIF-2 (на прикладі регуляції синтезу гемоглобіну) був розглянутий в главі 21.
Контроль вступу клітин до мітозу
Реплікація ДНК у еукаріотів відбувається протягом S-фази клітинного циклу, а мітоз, тобто розподіл подвоєного хромосомного матеріалу між дочірніми клітинами, починається тільки після підготовчої G2-фази. Вступу в М-фазу передує складна перебудова клітинної архітектури, що полягає в конденсації хроматину, розщепленні ядерної мембрани, реорганізації цитоскелета з формуванням мітотичного веретена тощо.
Ключовою подією в переході клітини до М-фази є активація специфічної протеїнкінази (сdc-білка), яка, після взаємодії з білком цикліном утворює каталітично активний комплекс, що фосфорилює численні клітинні білки, необхідні для реалізації мітотичного процесу.
Послідовність біохімічних реакцій включення мітозу є такою:
1. Утворення комплексу кінази cdc2 з цикліном.
Сdc2-кіназа — фермент з м.м. 34 кД; назва походить від гена, що кодує цей білок— cell-division-cycle gene (ген клітинного поділу — англ.).
Циклін — білок з м.м. 45 кД, концентрація якого поступово зростає протягом інтерфази і різко падає в кінці мітозу.
2. Фосфорилювання cdc2-кінази, що знаходиться в комплексі з цикліном; фосфорилювання відбувається за залишками Tyr-15 та Thr-161 ферменту. Двічі фосфорильована кіназа є неактивною, але готовою до включення своєї каталітичної активності.
3. Дефосфорилювання в молекулі cdc2-кінази залишку Tyr-15 з утворенням молекулярної форми ферменту, що фосфорильована тільки за Thr-161 і є каталітично активною.
Це дефосфорилювання спричиняється специфічною протеїнфосфатазою (білком cdc25) і є пусковою реакцією у всьому каскаді біохімічних реакцій, що реалізують перехід клітини до М-фази. Активація cdc25-фосфатази починається після завершення синтезу ДНК і відбувається протягом усього часу підготовки клітини до мітозу.
4. Каталітично активна cdc2-кіназа спричиняє фосфорилювання клітинних білків, які беруть участь у запуску мітотичної фази: ламінінів ядерної мембрани, що призводить до її розщеплення; білків мікротрубочок, що формують мітотичне веретено; Н1-гістонів, фосфорилювання яких веде до конденсації ядерного хроматину.
Викликає значний інтерес, що cdc2-кіназа включає ферментативний механізм, який призводить до обмеження її власної каталітичної активності і, як наслідок, виключення мітозу.
Обмеження активності cdc2-кінази досягається за рахунок розщеплення зв’язаного з нею цикліну. Молекулярний комплекс «циклін — монофосфорильована (за Thr-161) сdc2-кіназа» є субстратом для взаємодії з убіквітином(ubiquitin — англ.) — низькомолекулярним (8,5 кД) протеїном, що є універсальним лігандом для «мічення» білків, які підлягають подальшій деструкції під дією протеаз. Сполучення убіквітину з комплексом «циклін-сйс2-кіназа» призводить до обмеженого протеолізу цикліну і припинення клітинних реакцій, що реалізують процес мітозу.
Реакції біохімічного циклу, який контролює вступ еукаріотичної клітини у фазу мітозу, подані на схемі (рис. 22.13).
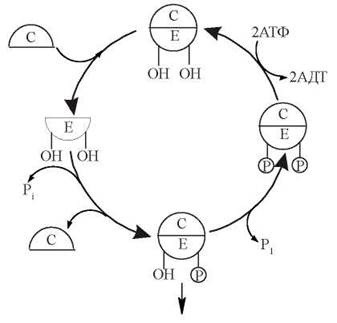
Рис. 22.13. Схема активації білків ініціації мітозу комплексом cdc-2-кінази (Е) з цикліном (С).