Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ IV. МОЛЕКУЛЯРНІ МЕХАНІЗМИ СПАДКОВОСТІ ТА РЕАЛІЗАЦІЇ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ
ГЛАВА 22. РЕГУЛЯЦІЯ ЕКСПРЕСІЇ ГЕНІВ. ГЕНЕТИЧНІ РЕКОМБІНАЦІЇ
Регуляція експресії генів є одним із фундаментальних механізмів адаптації живих організмів до умов навколишнього середовища, що змінюються. Враховуючи принципово різний ступінь складності та молекулярної організації геному у без’ядерних прокаріотів та вищих ядерних організмів — еукаріотів, механізми експресії генів у них значно відмінні.
22.1. РЕГУЛЯЦІЯ ЕКСПРЕСІЇ ГЕНІВ У ПРОКАРІОТІВ
Процес регуляції експресії генів у прокаріотів полягає у специфічній взаємодії певних білкових регуляторів з різними ділянками ДНК, що розміщені поряд із сайтами ініціації транскрипції. Такі взаємодії супроводжуються позитивним (індукуючим, активуючим) або негативним (гальмуючим, репресивним) впливом на рівень транскрипції, що, зрештою, впливає на швидкість синтезу відповідних ферментних та структурних білків.
Сучасна теорія регуляції експресії генів у прокаріотів була запропонована в 1961 р. французькими дослідниками Ф. Жакобом та Ж. Моно (F. Jacob, J. Monod) на підставі вивчення механізмів регуляції процесів транскрипції у E. Coli — улюбленого об’єкта молекулярної генетики.

Рис. 22.1. Жакоб (Jakob) Франсуа (народ.1920 р.), французький мікробіолог та генетик, професор кафедри генетики клітини в Колеж де Франс. Нобелівська премія (1965).

Рис. 22.2. Моно (Monod) Жак (1910-1976), французький біохімік та мікробіолог, професор Паризького ун-ту, Колеж де Франс. Нобелівська премія (1965).
Ф. Жакобом та Ж. Моно вивчалися молекулярно-генетичні механізми контролю в клітинах E.Coli біосинтезу β-галактозидази — ферменту, який розщеплює дисахарид лактозу на поживні моносахариди глюкозу та галактозу, що далі (переважно глюкоза) використовуються бактеріями як джерело вуглецю та метаболічної енергії:
![]()
При достатньому постачанні бактеріальних клітин глюкозою синтез β-галактозидази практично не відбувається, і в клітині міститься всього декілька молекул ферменту; після вичерпання глюкози як енергетичного субстрату додавання в культуральне середовище лактози спричиняє (індукує) різкий спалах синтезу β-галактозидази та ще двох ферментів, що беруть участь у метаболізмі лактози: β-галактозидпермеази та β-галактозидтрансацетилази.
Для пояснення механізмів контролю функції генів у E. Coli, які відповідають за метаболізм в клітинах енергетичного субстрату дисахариду лактози, Ф. Жакоб та Ж. Моно висунули теорію оперона(operon — працюю, дію; лат.), яка набула загальнобіологічного значення як модель, що розкриває молекулярні механізми індукції та репресії білкового синтезу на етапі транскрипції.
Оперон — комплекс генетичних елементів, що відповідає за координований синтез групи функціонально зв’язаних ферментних білків. Так, зокрема, у разі Lac-оперона E. Coli це — ферменти метаболізму лактози: β-галактозидаза, β-галактозидпермеаза та β-галактозидтрансацетилаза; His-оперон містить гени, що контролюють експресію дев’яти ферментів, які каталізують синтез необхідної для бактеріальної клітини амінокислоти гістидину тощо.
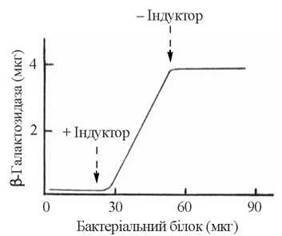
Рис. 22.3. Індукція синтезу β-галактозидази у E. Coli в умовах внесення в інкубаційне середовище лактози. Концентрація бактеріального білка (вісь абсцис) відображує активність поділу клітин у культурі.
До складу оперона входять (рис. 22.4):
(1) структурні гени(Z,Y,A), що містять інформацію відносно первинної структури поліпептидів, які транскрибуються з даного оперона; в Lac-опероні E. Coli — це гени зазначених вище трьох ферментів метаболізму лактози;
(2) контрольні сайти, до яких належать:
а) промотор(p) — ділянка ДНК, що первинно взаємодіє з РНК-полімеразою; в промоторах E. Coli міститься також особлива ділянка, з якою взаємодіє білок-активатор катаболітних генів (САР-білок — catabolite gene activator protein, англ.), що контролює зв’язування РНК-полімерази з промотором;
б) оператор (o) — ділянка ДНК, з якою може специфічно зв’язуватися білок-репресор. Оператор безпосередньо прилягає до структурних генів, і його зв’язування з репресором протидіє зчитуванню РНК-полімеразою інформації із структурних генів.
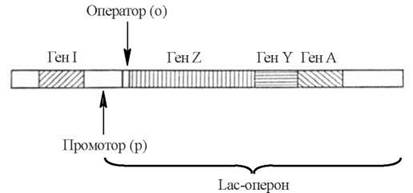
Рис. 22.4. Схематична будова Lac-оперону E. Coli.
Регуляторний ген (I), експресія якого призводить до продукування білкових регуляторів (репресорів), що блокують зчитування інформації із структурних генів оперону, не розглядається як інтегральна складова останнього, оскільки він може бути локалізований на певній відстані від оперону, функцію якого він контролює.
Схема діяльності Lac-оперону
Активність Lac-оперона є об’єктом подвійної — негативної та позитивної регуляції.
А. Репресія Lac-оперону.
У цьому стані синтез в-галактозидази та інших ферментів метаболізму лактози практично не відбувається — відповідні гени репресовані. Цей стан має місце в умовах присутності достатньої кількості глюкози; оскільки глюкоза є катаболітом лактози, стан гальмування синтезу ферментів метаболізму лактози в присутності глюкози отримав назву стану катаболітної репресії, хоча насправді глюкоза не є безпосереднім репресором, а впливає на синтез зазначених ферментів іншим, більш складним шляхом (див. нижче).
Блокування (репресія) структурних генів Lac-оперону здійснюється за рахунок продукування регуляторним геном спеціального білкового репресора, який, зв’язуючись із оператором (певною послідовністю нуклеотидів ДНК) блокує транскрипцію структурних генів РНК-полімеразою (рис. 22.5).
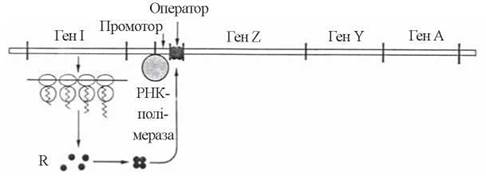
Рис. 22.5. Механізм репресії Lac-оперону.
Зв’язування репресора (R) з оператором протидіє просуванню РНК-полімерази впродовж оперону і транскрипції дистально розташованих структурних генів Z,Y,A.
Lac-репресор — це білок тетрамерної будови з м.м. 38 кД; сполучаючись з ДНК операторного локусу, який локалізований між промотором (з яким первинно зв’язується РНК-полімераза) та початком структурних генів, репресор протидіє зчитуванню інформації з генів Z,Y,A.
За розглянутим механізмом здійснюється негативна регуляція функції Lac-оперону E. Coli.
Б. Індукція Lac-оперону.
Це стан активного синтезу ферментів метаболізму лактози, що спостерігається за умов вичерпання глюкози як енергетичного субстрату та внесення в культуральне середовище лактози (рис. 22.3).
1. Одним із механізмів активованого синтезу ферментів метаболізму лактози в умовах, що розглядаються, є безпосередня взаємодія індуктора (лактози або деяких її структурних аналогів) з репресором (як вільним, цитоплазматичним, так і зв’язаним з операторним локусом).
Зв’язування індуктора з Lac-репресором спричиняє конформаційні зміни в молекулі останнього і втрату ним властивості взаємодіяти з ділянкою ДНК операторного локусу, що розгальмовує транскрипцію із структурних генів — рис. 22.6.
Найбільш активним фізіологічним індуктором Lac-оперону є алолактоза — дисахарид, що утворюється внутрішньоклітинно з лактози в реакції трансглікозилювання; cинтетичним індуктором є сполука ізопропілтіогалактозид.
2. Крім залежності від зняття індуктором репресорного блока, синтез ферментів Lac-оперону контролюється також особливим білком, що має назву білка-активатора катаболітних генів (CAP). Встановлено, що зв’язування РНК-полімерази з промотором і ініціація транскрипції можливі лише за умов утворення комплексу САР з циклічним аденозинмонофосфатом (3',5'-цАМФ, сАМР) та сполучення цього комплексу з певною ділянкою ДНК промотора (рис. 22.6).
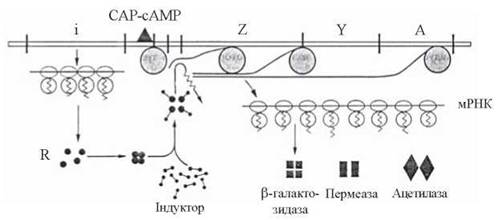
Рис. 22. 6. Механізм індукції Lac-оперону. Присутність індуктора інактивує репресор, який втрачає здатність до блокування гена-оператора; в разі інактивації репресора (а також за умов сполучення промотору з комплексом САР-сАМР) РНК-полімераза починає транскрибувати структурні гени.
Фізіологічне значення цього феномена полягає в можливості позитивного контролю зчитування інформації із структурних генів Lac-оперону під впливом 3',5'- цАМФ, концентрація якого в клітині, в свою чергу, визначається активністю метаболізму глюкози. Активація аденілатциклази, що призводить до генерації 3',5'- цАМФ, відбувається в умовах зменшення в культуральному середовищі джерел вуглецю (глюкози або гліцеролу), що через утворення комплексу САР-сАМР стимулює початок транскрипції РНК-полімеразою генів Z,Y,A.
Таким чином, при дії комплексу САР-сАМР реалізується механізм позитивної регуляції експресії генів Lac-оперону.