Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ V. ГОРМОНИ В СИСТЕМІ МІЖКЛІТИННОЇ ІНТЕГРАЦІЇ ФУНКЦІЙ ОРГАНІЗМУ
ГЛАВА 23. ГОРМОНАЛЬНА РЕГУЛЯЦІЯ МЕТАБОЛІЗМУ ТА БІОЛОГІЧНИХ ФУНКЦІЙ КЛІТИНИ. I. БІОХІМІЧНІ СИСТЕМИ ВНУТРІШНЬОКЛІТИННОЇ ТРАНСДУКЦІЇ ГОРМОНАЛЬНИХ СИГНАЛІВ
23.3. МОЛЕКУЛЯРНО-КЛІТИННІ МЕХАНІЗМИ ДІЇ СТЕРОЇДНИХ ТА ТИРЕОЇДНИХ ГОРМОНІВ
Гормони другої групи — стероїди (кортикостероїди, гормони статевих залоз, похідні вітамінів групи D), тиреоїдні гормони.
Подібно до гормонів білково-пептидної групи та біогенних амінів — похідних амінокислот, необхідною умовою здійснення біологічних ефектів гормонами цієї групи також є їх зв’язування в тканинах- мішенях із специфічними рецепторними молекулами. Справедливість висловлювання засновника вчення про клітинні рецептори Пауля Ерліха (P. Ehrlich) — ’Corpora non agun nisifixata» («Речовини не діють, якщо не фіксуються» — пер. з лат.) яскраво ілюструють експериментальні результати, подані на рис. 23.10.
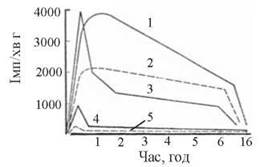
Рис. 23.10. Динаміка розподілу Н3-естрадіолу в тканинах щура-самиці (вісь абсцис — ступінь зв’язування гормону в тканині, імп./хв г тканини; вісь ординат — час після введення гормону, год) 1 — матка; 2 — vagina; 3 — печінка; 4 — кров; 5 — м’язи.
Але на відміну від гормонів — пептидів та біогенних амінів, рецептори для гормонів цієї групи локалізовані внутрішньоклітинно — в цитозолі, звідки гормонорецепторні комплекси транслокуються в ядро, де і взаємодіють з чутливими сайтами ДНК ядерного хроматину, спричиняючи активацію генів для відповідних ферментних білків. Тоді як гормони першої групи спричиняють активацію вже існуючих ферментних молекул, дія на клітини-мішені стероїдів та тиреоїдних гормонів призводить до стимуляції біосинтезу нових ферментних молекул за рахунок активації процесів транскрипції їх мРНК (рис. 23.11).
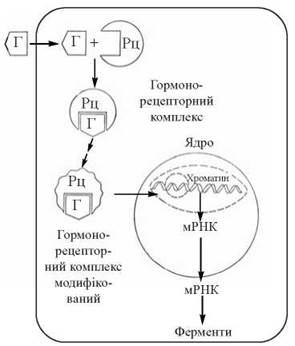
Рис. 23.11. Схема біохімічних механізмів дії стероїдних та тиреоїдних гормонів.
Згідно із зазначеним, білково-пептидні гормони та біогенні аміни контролюють процеси швидкої адаптації організму, які потребують термінового включення певного біохімічного процесу або фізіологічної функції (глікогенолізу, ліполізу, м’язового скорочення), тоді як біологічна дія гормонів другої групи повільніша (потребує для свого проявлення декількох годин), вони відповідають за процеси довготривалої адаптації організму.
Будова стероїдних та тиреоїдних рецепторів
Нетрансформовані взаємодією з гормоном рецептори є димерними білками з молекулярною масою, що коливається від 50 до 190 кД, і константами седиментації від 3,5 до 4,5 s (табл. 23.2).
Таблиця 23.2. Фізико-хімічні властивості цитозольних рецепторів стероїдних гормонів (В.Б. Розен, 1984)
Рецептори гормонів |
М. м, кД |
Константа седиментації, s |
Естрогенів |
80 - 120 |
4,0 - 4,5 |
Андрогенів |
80 - 100 |
3,5 - 4,0 |
Прогестагенів |
50 - 100 |
3,6 - 4,0 |
Глюкокортикоїдів |
70 - 190 |
3,5 - 4,0 |
Альдостерону |
60 - 150 |
4,0 - 4,5 |
Молекулярні механізми дії
Послідовність клітинних та біохімічних реакцій, за рахунок якої стероїдні та тиреоїдні гормони реалізують свої біологічні ефекти, має вигляд:
проникнення гормону всередину клітини —► сполучення гормону з цитозольним рецептором —► модифікація (активація) рецептора у складі гормоно-рецепторного комплексу —► транслокаціямодифікованого гормонорецепторного комплексу в ядро —► взаємодія комплексу зі специфічною ділянкою ДНК хроматину —► активація специфічних генів —► транскрипція мРНК —► синтезферментних білків, що реалізують біологічні ефекти гормону.
Взаємодія рецепторів стероїдних та тиреоїдних гормонів із своїми специфічними лігандами зумовлена їх молекулярною будовою, що складається з окремих структурно-функціональних доменів, які відрізняються первинною структурою та конформацією (рис. 23.12):
- домену, що сполучається з гормоном (складається з 240 амінокислотних залишків);
- домену, що сполучається з певними послідовностями ДНК (складається з 66 амінокислотних залишків);
- домену, що зв’язується з додатковими регуляторами транскрипції (N-термінальний домен).
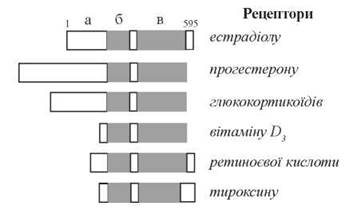
Рис. 23.12. Доменна організація білкових рецепторів гормонів: а — домен, що зв’язує регулятори транскрипції; б — домен, що взаємодіє з ДНК; в — домен, що зв’язує гормон.
Взаємодія білкових рецепторів гормонів — активаторів транскрипції з ДНК відбувається в певних місцях промоторних ділянок геному, що знаходяться «зліва» від сайтів ініціації транскрипції(приблизно, в регіоні «250 нуклеотидів») і регулюють експресію розташованих на відстані генів («справа» від промотора та сайта ініціації транскрипції «+1») (рис. 23.13).
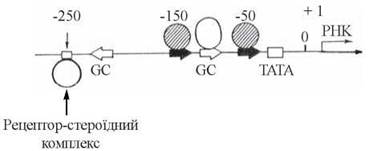
Рис. 23.13. Ділянки промотору, з якими взаємодіють стероїд-рецепторні комплекси (сайт-250) та інші білки-активатори транскрипції (стресорні білки, активатори синтезу металотіонеїну).
Здатність домена гормонального рецептора взаємодіяти з певними ділянками ДНК визначається особливостями структури як рецептора, так і відповідних сайтів ДНК. Ці рецепторчутливі ділянки ДНК мають структуру паліндромів і складаються із специфічних (для кожного рецептора) нуклеотидних послідовностей з симетричних 6 пар нуклеотидів, розташованих зліва та справа від проміжної 3-нуклеотидної послідовності (-NNN-) — «спейсера»:

У взаємодії активованих гормонами стероїдних та тиреоїдних рецепторів зі специфічними ділянками ДНК беруть участь також певні ділянки рецепторних білків, що мають будову цинкових пальців та глобулярних Zn-вмісних доменів (рис. 23.14) — унікальних просторових утворень, які властиві всім різновидам білків, що виступають як регулятори транскрипції (гормональним рецепторам, білкам — активаторам синтезу металотіонеїну тощо).
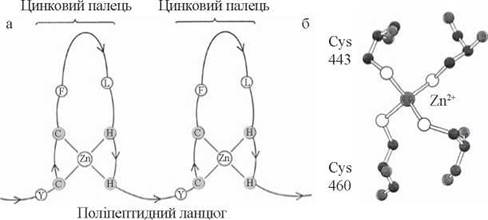
Рис. 23.14. Утворення цинкових пальців у білках (а) та будова Zn-вмісного домену глюкокортикоїдного рецептора, що розпізнає специфічну паліндромну послідовність у молекулі ДНК (б).
Молекулярна модель зв’язування Zn-вмісних доменів димерного рецептора глюкокортикоїдів з паліндромними ділянками молекули ДНК подана на рис. 23.15.
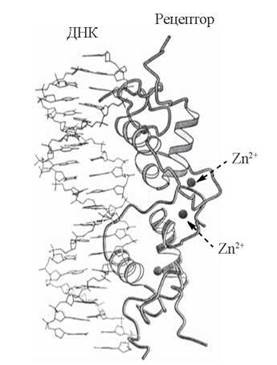
Рис. 23.15. Взаємодія димерного білка — рецептора глюкокортикоїдів із специфічними сайтами ДНК.