Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ V. ГОРМОНИ В СИСТЕМІ МІЖКЛІТИННОЇ ІНТЕГРАЦІЇ ФУНКЦІЙ ОРГАНІЗМУ
ГЛАВА 24. ГОРМОНАЛЬНА РЕГУЛЯЦІЯ МЕТАБОЛІЗМУ ТА БІОЛОГІЧНИХ ФУНКЦІЙ КЛІТИНИ. II. ГОРМОНИ - ПОХІДНІ ПЕПТИДІВ ТА АМІНОКИСЛОТ
24.3. ТИРЕОЇДНІ ГОРМОНИ
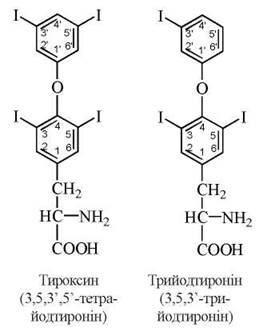
Тиреоїдні гормони є істинними гормонами, що синтезуються в спеціалізованих епітеліальних клітинах фолікулів щитовидної залози — тиреоцитах. До цієї групи належать похідні амінокислоти L-тирозину — 3,5,3', 5'-тетрайодтиронін (тироксин) та 3,5,3'-трийодтиронін:
Біосинтез тиреоїдних гормонів потребує наявності йодидів, що вибірково накопичуються в щитовидній залозі, яка поглинає їх із крові; із загальних 20-30 мг йоду, що містяться в тілі людини, 10-15 мг акумулюється в колоїді фолікулів щитовидної залози.
Біосинтез тиреоїдних гормонів складається з таких етапів (рис. 24.5):
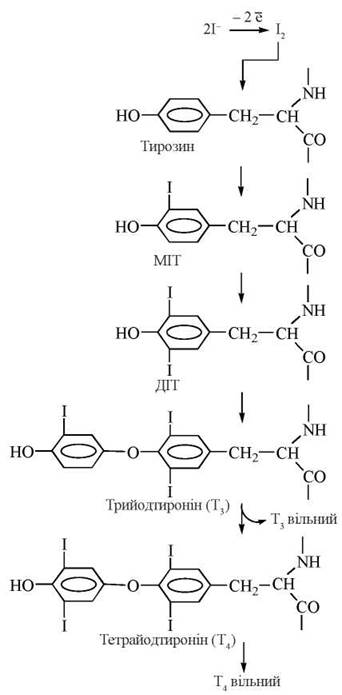
Рис. 24.5. Схема біосинтезу тиреоїдних гормонів.
1) акумуляція щитовидною залозою йодидів крові (І) за допомогою «йодидного насосу» та їх окислення йодидпероксидазою до молекулярного йоду:
![]()
2) синтез специфічного білка колоїду щитовидної залози тиреоглобуліну та йодування його тирозинових залишків з утворенням монойодтирозинів (МІТ) та дийодтирозинів (ДІТ). Тиреоглобулін — глікопротеїн з м.м. 660 кД, що має олігомерну структуру (складається з двох субодиниць); у складі білка — 115 залишків тирозину, 15-20 % яких підлягають йодуванню;
3) перетворення йодованих тирозилових залишків на йодовані тироніни (у складі молекули тиреоглобуліну);
4) секреція йодованого тиреоглобуліну в порожнину фолікула, де цей високомолекулярний попередник тиреоїдних гормонів зберігається у складі колоїду;
5) поглинання (за умов фізіологічних потреб та стимуляції тиреотропіном) йодованого білка тиреоцитами, включення його в фаголізосоми, розщеплення молекули йодованого тиреоглобуліну(лізосомальними тиреокатепсинами) з утворенням вільних молекул трийодтироніну (Т3) та тетрайодтироніну (Т4) і вихід їх через базальні мембрани клітин у кров.
Із двох йодованих тиронінів більш активним є Т3, специфічна гормональна активність якого перевищує відповідну активність Т4 у 4-5 разів; у периферійних тканинах більша частина Т4перетворюється на Т3, і сумарна біологічна дія тиреоїдних гормонів в організмі на 90-95 % забезпечується саме Т3.
Біологічні функції тиреоїдних гормонів — полягають у контролі процесів енергетичного обміну, біосинтезу білка та морфогенезу:
1. Стимуляція біоенергетичних процесів у тканинах при дії гормонів щитовидної залози позначається збільшенням швидкості тканинного дихання (поглинання О2), активності мітохондріальних ферментів елетронотранспортного ланцюга, підвищенням рівня катаболізму вуглеводів, ліпідів, амінокислот. Разом з тим, збільшення поглинання тканинами кисню в умовах дії тиреоїдних гормонів не супроводжується адекватним зростанням фосфорилювання АДФ до АТФ, в результаті чого коефіцієнт окисного фосфорилювання (Рi/О) зменшується, тобто відбувається роз’єднання дихання та окисного фосфорилювання.
2. Тиреоїдні гормони є потужними стимуляторами процесів морфогенезу, нео- натального закладення, формування і розвитку тканинних структур та органів; уроджений гіпотиреоз у людини супроводжується важкою затримкою психічного розвитку новонароджених дітей. В основі позитивного впливу гормонів щитовидної залози на морфогенез та розвиток тканин лежить їх специфічний активуючий вплив на процеси транскрипції генів, що контролюють зазначені процеси.
Молекулярні механізми дії
Рецептори тиреоїдних гормонів локалізовані в ядрах клітин-мішеней. У взаємодії з гормонами беруть участь специфічні рецептори α-1 та β-1, які є білками з м.м. 47 і 52 кД, відповідно. Спорідненість цих рецепторів до Т3 значно більша, ніж до Т4.
Молекули тиреоїдних рецепторів мають три структурних домени: N-термінальний домен; центральний ДНК-зв’язуючий домен з двома цинковими пальцями; С-термінальний домен, що сполучається з гормоном (глава 23). Утворення комплексу гормон-рецептор призводить до конформаційних змін у молекулі рецептора; при цьому ДНК-зв’язуючий домен рецептора стає спроможним до взаємодії із специфічними сайтами на промоторах генів, які контролюють транскрипцію мРНК та синтез білків, що відповідають за прояви біологічних ефектів тиреоїдних гормонів.
Патологія щитовидної залози
Численні клінічні форми патологічних процесів, що стосуються щитовидної залози, призводять у кінцевому підсумку до зменшення або збільшення її функціональної активності (гіпотиреоз або гіпертиреоз, відповідно).
а) Гіпотиреоз — патологічний стан, який розвивається внаслідок дефіциту в організмі вільних Т3 та Т4 або неадекватної реакції тканин-мішеней на дію гормонів. Така ситуація може бути спричиненою дефектами синтезу тиреоїдних гормонів на різних його стадіях внаслідок:
- порушення акумуляції йодидів залозою (внаслідок йодного дефіциту або неспроможності залози накопичувати мікроелемент);
- порушень у ферментних системах, що використовують йодиди для утворення гормонально активних молекул Т3 та Т4;
- порушень у рецепторних та трансдукуючих системах, які трансформують гормональний сигнал в специфічні ефекти тиреоїдних гормонів.
Поширеною формою гіпотиреозу є ендемічний зоб — хвороба, яка часто зустрічається в місцевостях, де наявний екзогенний дефіцит йоду в питній води та продуктах харчування. Захворювання характеризується збільшенням розміру залози — «зобом», спричиненим компенсаторною активацією виділення ТТГ; специфічними проявами хвороби ємікседема (слизистий набряк), загальмованість нейро-психічних процесів, рухова млявість, апатичність, у разі прояву патології в ранньому дитячому віці — значна затримка розумового та фізичного розвитку.
б) Гіпертиреоз — патологічний стан, що пов’язаний із надлишковим утворенням залозою тиреоїдних гормонів. Різні за механізмами походження клінічні форми гіпертиреозу (базедова хвороба, хвороба Грейвса тощо) супроводжуються тиреотоксикозом («токсичний зоб»), характерними проявами якого є збільшення основного обміну (поглинання організмом кисню в стані спокою), підвищена збудливість нервової системи, психічна дратівливість, тахікардія, схуднення хворих внаслідок переважання катаболічних процесів.