Практическая химия белка - А. Дарбре 1989
Определение состава белковых олигомеров. Получение мономеров и полипептидных цепей
Сшивание олигомера и мономера
Дисульфидные связи
О факте сшивания отдельных субъединиц свидетельствуют данные анализа N-концевых аминокислотных остатков. Например, выход N-концевой аминокислоты в количестве >1 моль/моль белка может служить доказательством присутствия в олигомере двух идентичных полипептидных цепей. Аналогичным образом выход N-концевых аминокислот (исключая серин) на уровне 50% может служить признаком того, что N-концевая аминокислота одной из двух цепей блокирована. Поскольку выход N-концевой аминокислоты зависит от многих факторов, в истолковании полученных результатов следует избегать поспешных выводов.
В белках были найдены различные типы поперечных связей: дисульфидная, альдольная и альдиминная. Наиболее лабильные из них — S—S-связи, поэтому на практике чаще всего прибегают именно к расщеплению дисульфидных связей. Для разделения образующихся полипептидных цепей используют обычные методы, принятые при разделении мономеров (разд. 1.3.2).
Дисульфидные связи расщепляют путем окисления, восстановления, с помощью реакций нуклеофильного замещения (HSO-3, CN-, ВН-4, Н-);[40, 106] (см. также гл. 2). При окислении S—S-связей образуются два остатка цистеиновой кислоты [160]. Дополнительные сульфогруппы придают молекуле белка заметную гидрофильность, повышая его растворимость в воде, в особенности в области низких pH. Однако окисление дисульфидных связей сопровождается модификацией других аминокислот и частичным гидролизом лабильных пептидных связен. Поэтому для введения сульфогруппы предпочитают проводить восстановительное расщепление дисульфидных связей, а затем мягкое окисление SH-групп реагентами ряда сультонов (внутренних сложных эфиров сульфокислот) (см. разд. 1.5.1.2 и 2.2.2.3).
Обычно дисульфидные связи расщепляют восстановлением, полноту реакции контролируют с помощью реактива Эллмана [69, 102]. После восстановления образующиеся SH-группы алкилируют с целью исключить окисление с образованием цистина. Алкилирующий агент выбирают в соответствии с поставленной дополнительной задачей, например можно стремиться придать белку более высокую растворимость в водном буфере, ввести дополнительные точки для гидролиза трипсином или ввести лабильную защитную группу в расчете на последующую регенерацию SH-групп. Иногда защита по SH-гpyппам может оказаться излишней, поскольку белок с восстановленными S—S-связями может быть устойчив в уксусной кислоте на холоду при pH 3 [7].
1.5.1.1. Расщепление дисульфидов.
Окисление цистеина или дисульфидных связей в белках рекомендуется проводить с помощью надмуравьиной кислоты [78] (см. также разд. 2.2.3.1).
После проведения окисления определяют аминокислотный состав исследуемого белка, контролируя прежде всего содержание цистеиновой кислоты. Выход цистеиновой кислоты должен составлять ≥85%, а выход S,S-диоксида метионина (при наличии остатка метионина) не менее 95%; триптофан при окислении разрушается полностью. При наличии хлоридов может происходить модификация тирозина, поэтому перед анализом рекомендуется удалять из образца ионы хлора. Колонку с дауэксом 2 (100 меш) в ацетатной форме промывают З М ацетатом натрия до отсутствия в элюате ионов хлора (контроль — реакция с нитратом серебра). Затем через колонку пропускают раствор белка (0,1 г белка на 1 мл объема ионита) и промывают небольшим объемом буфера. При этом следует учитывать возможность потери белка на колонке.
Восстановление S—S-связей проводят главным образом 2-меркаптоэтанолом. Однако при недостаточно высоком избытке 2-меркаптоэтанол образует смешанные дисульфиды, к тому же он обладает неприятным запахом. Удовлетворительными свойствами обладают дитиотреит и дитиоэритрит, которые при небольшом избытке обеспечивают полноту восстановления. В ряде работ дисульфидные связи восстанавливали трибутилфосфином [150—152] (более детально эти методы обсуждаются в разд. 2.2.1).
Дисульфидные связи, соединяющие две или более полипептидные цепи, обычно доступны действию растворителей или реагентов даже в отсутствие денатурирующих агентов, тогда как внутрицепочечные S—S-связи чаще всего бывают маскированы и недоступны. Это обстоятельство позволяет проводить избирательное восстановление «экспонированных» S—S-связей. Полное восстановление дисульфидов рекомендуют проводить в слабокислой среде в присутствии мочевины или гунидингидрохлорида. Ни один из денатурирующих агентов не свободен от недостатков. Например, примеси мочевины в образце генерируют цианат-ионы на стадии конденсации при деградации по Эдману, что может препятствовать секвенированию. Опыт применения гуанидин-HCl не столь поучителен, однако при использовании хорошо очищенного препарата проблем при секвенировании не наблюдалось.
При исследовании уG-иммуноглобулина был разработан метод восстановления межцепочечных S—S-связей при сохранении внутрицепочечных [53].
В полученных фрагментах определяют содержание SH- групп или же смесь фракционируют в присутствии восстанавливающих агентов или в кислой среде. Для удобства работы тиолсодержащие фрагменты переводят в более стабильные или более растворимые производные.
Наиболее изученным, но вместе с тем не свободным от недостатков способом защиты SH-групп является карбоксиметилирование [68]. Следует отметить, что, если стадию конверсии тиазолинонов в фенилтиогидантоины при секвенировании по Эдману проводить в недостаточно мягких условиях, наблюдаются значительные потери S-карбоксиметилцистеина [84]. Полезным приемом является алкилирование 14С-иодоацетамидом, однако с успехом применяются и другие реагенты.
Если в дальнейшем планируется проводить исследование по ренатурации белка, можно получать производные с лабильными защитными группами. Некоторые из рекомендованных реагентов требуют осторожности в обращении. Например, иодоуксусная кислота — аллерген, и, как правило, с этим реактивом работают в резиновых перчатках при хорошей вентиляции (см. также гл. 2).
1.5.1.2. Обратимая модификация. При действии на цистин сульфита образуются цистеин и цистеинсульфонат (сульфоцистеин) (см. также разд. 2.2.4).
![]()
Дисульфидные связи можно реконструировать путем восстановления модифицированного белка и его ренатурации в присутствии кислорода воздуха. Окислительный сульфитолиз дисульфидов в присутствии солей меди приводит к образованию S-сульфонатов [37].
![]()
Аналогичные результаты были получены при реакции белка с сульфитом натрия в 8 М мочевине в присутствии кислорода воздуха и следов цистеина [31]. Восстановление цистина дитиотреитом и последующая обработка тетратионатом натрия приводит к образованию S-сульфоцистеина с количественным выходом [85]. S-сульфоцистеин устойчив при нейтральном pH и в условиях деградации по Эдману, однако при восстановлении образует цистеин. При гидролизе белков 6М НСl S-сульфонистеин превращается в цистин [37]. Серьезную проблему представляет контроль за полнотой реакции: при использовании [35S]сульфита контроль ведут по включению метки [31].
При взаимодействии w-толуолсультона с восстановленным белком при pH 8,3 образуется S-2-сульфобензилцистеии [153].
![]()
Обратную реакцию проводят путем восстановления натрием в жидком аммиаке; с целью воспрепятствовать расщеплению связей Тrе-Рrо в реакционную смесь добавляют амид натрия. Белки, модифицированные путем сульфитолиза и сульфобензилирования, хорошо раcтворимы в воде.
Окислительный сульфитолиз [31]. Метод с успехом применялся при исследовании альдолазы, лектатдегидрогеназы и пепсиногена. Полнота реакции достигается только в диапазоне pH 7,0—8,5, что свидетельствует об участии в реакции протонированной аминогруппы цистеина. Образец белка (2 мг/мл) инкубируют в присутствии кислорода воздуха при 25 °С в течение 1 ч в растворе следующего состава: 0,1 М трис-НСl (pH 8,4)+8 М мочевина (или 6 М гуанидин-НСl)+0,05 М сульфит натрия, 0,2 мМ цистеин. По завершении реакции реакционную смесь диализуют и высушивают лиофильно.
После сульфитолиза активный фермент (альзолазу) получают по следующей методике. Образец белка (0,5—1 мг/мл) растворяют в 0,1 М трис-НСl (pH 7,5), содержащем 4 М мочевину, добавляют 2-меркаптоэтанол (до концентрации 0,7 моль/л) и инкубируют при 25 °С в течение 2 ч. Затем реакционную смесь разбавляют в 10 раз буферным раствором следующего состава: 0,1 М трис-НС1 (pH 7,5)+2,5 мг/мл бычий сывороточный альбумин+25 мМ ЭДТА+10 мМ 2-меркаптоэтанол; инкубируют в присутствии кислорода воздуха и спустя 15 мин определяют активность ренатурированного фермента.
S-Сульфобензилирование [153]. Образец белка (100 мкмоль) в 100 мл 0,5 М бикарбоната натрия в смеси с пропанолом (1:1) инкубируют в атмосфере азота с трибутилфосфином (330 мкмоль) и w-толуолсультоном (204 мг, 1200 мкмоль). Избыток w-толуолсультона отделяют фильтрованием, фильтрат упаривают до остаточного объема 40 мл и диализуют против воды. На аминокислотном анализаторе S-сульфобензилцистеин элюируется непосредственно после цистеиновой кислоты. При проведении реакции с инсулином выход производных цистеина количественный.
Снятие S-сульфобензильной группы [90]. 300 мг белка тщательно высушивают, растворяют в безводном жидком аммиаке (300 мл) в круглодонной колбе при охлаждении сухим льдом в ацетоне. Для защиты лабильных пептидных связей остатка пролива в реакционную смесь добавляют ~120 мг амида натрия. Последующие стадии проводят при температуре кипения жидкого аммиака. Предварительно приготовленный раствор металлического натрия в жидком аммиаке добавляют по каплям до тех пор, пока реакционная смесь не приобретает слабую голубую окраску. Выдерживают 30 с и добавляют несколько капель (2—3) ледяной уксусной кислоты до исчезновения окраски, затем аммиак упаривают до остаточного объема 10 мл. Остаток упаривают в вакууме на роторном испарителе и высушивают досуха на водоструйном насосе.
1.5.1.3. Окисление. Реконструкция большинства дисульфидных связей в некоторых белках достигается при окислении кислородом воздуха, т. е. при аэрировании раствора восстановленного белка [70]. По-видимому, молекуле нативного белка свойственна термодинамически наиболее выгодная конформация, и, следовательно, главным фактором, способствующим «правильному» сближению тиольных групп, является кооперативное (согласованнное) взаимодействие функциональных групп полипептидной цепи. Если распределение дисульфидных связей в белке определяется главным образом конформацией белка-предшественника (например, в случае инсулина проинсулином), то такие белки ренатурируют с трудом. Далее приводится общепринятая методика окисления белков с целью ренатурации, однако оптимальные условия реакции определяются в зависимости от индивидуальных свойств исследуемого белка, его первичной структуры и трехмерной укладки полипептидной цепи. Например, добавление 2-меркаптоэтанола и увеличение температуры до 38 °С оказались полезными при ренатурации лизоцима, но неэффективными в случае рибонуклеази [56]. В случае других белков оптимальную концентрацию 2-меркаптоэтанола подбирают эмпирическим путем. Оба фактора — низкая концентрация белка и присутствие 2-меркаптоэтанола — препятствуют агрегации субъединиц и образованию межмолекулярных дисульфидных связей. Предполагают, что 2-меркаптоэтанол катализирует дисульфидный обмен, необходимый для коррекции неправильно замкнутых остатков цистеина.
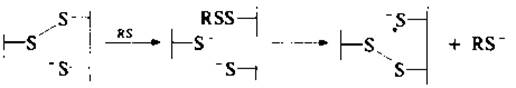
Ионы металлов из-за связывания с тиолами ингибируют активацию. При активации используют медленный диализ денатурирующих агентов, например мочевины. Для доступа кислорода вполне достаточно проводить реакцию в открытых сосудах; активная аэрация не рекомендуется, так как может вызвать денатурацию белка.
Методика 1 [56]. Ренатурацию восстановленного лизоцима (0,025 мг/мл) проводят в 0,1 М трис-буфере (pH 8,5) при 38 °С в течение 2 ч в присутствии 2-меркаптоэтанола (при молярном отношении 2-меркаптоэтапол : цистеин = 180 : 1). Выход активированного продукта 70%.
Методика 2 [52]. Восстановленные цепи инсулина (5 мг белка/мл) осаждают при pH 3,8 и тщательно промывают деаэрированным буфером на холоду. Окисление кислородом воздуха проводят в глициновом буфере (pH 10,6) при 35 °С. Ренатурированный белок имеет 50%-ную активность по сравнению с кристаллическим препаратом. Активность полученного гормона зависит от относительного содержания в реакционной смеси полипептидных цепей; максимальный выход достигается при соотношении А: В=1,5:1.