БИОХИМИЯ И МОЛЕКУЛЯРНАЯ БИОЛОГИЯ - В. ЭЛЛИОТ - 2002
ГЛАВА 19. ДНК: СТРУКТУРА И ЛОКАЛИЗАЦИЯ В КЛЕТКАХ
В этой главе мы познакомимся со структурой ДНК и понятием гена, а в двух последующих будет рассказано о синтезе ДНК и о ее роли в образовании белка. Главная функция ДНК связана с ее участием в синтезе белков, аминокислотная последовательность которых определяет все множество процессов, составляющих основу жизни в течение миллионов лет эволюции.
Что такое нуклеиновые кислоты?
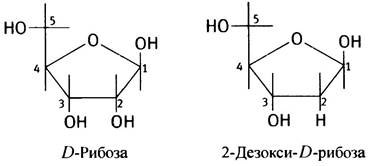
Дезоксирибонуклеиновая кислота, или ДНК, впервые была выделена из клеточных ядер. Поэтому она была названа нуклеиновой (от лат. nucleus - ядро). Ее кислотные свойства обусловлены присутствием в составе молекулы остатков фосфорной кислоты. В качестве углеводного компонента выступает 2-дезокси-D-рибоза. В совокупности все это и определило название - дезоксирибонуклеиновая кислота. В клетках присутствует и другой тип нуклеиновых кислот - рибонуклеиновые, в которых углевод представлен D-рибозой. Поэтому и называются они рибонуклеиновыми, или РНК (см. главу 21). В отличие от D-рибозы 2-дезокси-D-рибоза не содержит кислорода при втором углеродном атоме; ее обычно называют просто дезоксирибозой.
В эукариотических клетках основная масса ДНК заключена в ядре (незначительные количества присутствуют в митохондриях и хлоропластах), в то время как большая часть РНК находится в цитоплазме, хотя синтезируется она в ядре (кроме РНК митохондрий и хлоропластов). Хотя синтез нуклеотидов был уже рассмотрен в главе 18, для удобства изложения мы вновь вернемся к этому классу соединений. Но сначала о ДНК.
Первичная структура ДНК
ДНК - полинуклеотид. Строение нуклеотида можно представить следующим образом: фосфат-сахар-основание.
А так выглядит структура дезоксирибонуклеотида, для простоты приведенная в неионизированной форме.
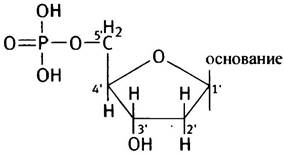
Сахар представлен, как уже отмечалось выше, 2-дезокси-D-рибозой (дезоксирибозой) и находится в форме фуранозы, образуя пятичленный цикл. При нумерации атомов дезоксирибозного цикла добавляется штрих ('), чтобы отличить ее от нумерации атомов в основаниях, поэтому атомы углерода в углеводном остатке обозначаются как 1', 2', 3', 4' и 5' (произносится «пять штрих» и т. д.) и номера записываются с внешней стороны цикла.
Какие основания входят в ДНК?
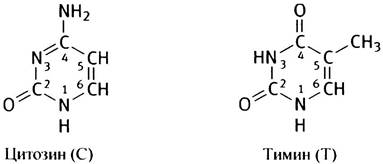
В отличие от белков, образованных 20 аминокислотами, в составе ДНК встречается только 4 различных основания, структура которых ненамного сложнее, чем структура аминокислот. Основания представлены аденином, гуанином, цитозином и тимином, или, сокращенно, А, G, С и Т соответственно. А и G - пурины, С и Т - пиримидины. Нумерация атомов в основаниях записывается внутри цикла.
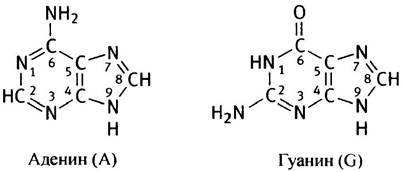
Обратите внимание на метильную группу в тимине! В дальнейшем названия оснований будут приводиться в сокращенном варианте.
Основания могут существовать в таутомерных формах (кето-енол и амино-имино), и при нейтральных значениях pH основания в ДНК находятся преимущественно в форме кето- и амино-таутомеров:
![]()
а не в енольной и иминной формах:
![]()
Связь оснований с дезоксирибозой
Первый атом углерода сахара связывается гликозидной связью с атомом азота в положении 9 или 1 пуринового либо пиримидинового основания соответственно. Эта связь называется β-гликозидной, что определяется структурой углеводного кольца.
Соединение, состоящее из основания и углевода, называется нуклеозидом; если углевод представлен дезоксирибозой -дезоксирибонуклеозидом. Все дезоксирибонуклеозиды имеют свои названия.
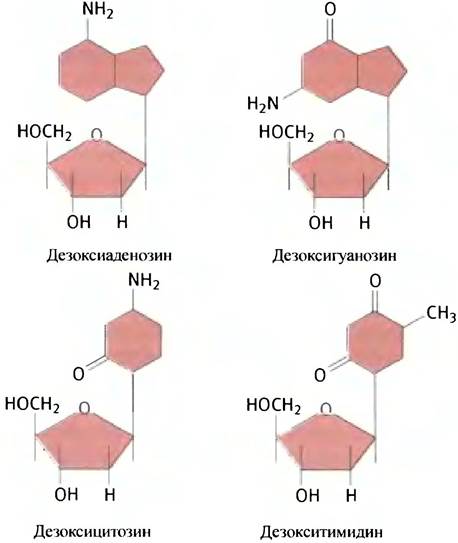
Нуклеозид, содержащий в качестве основания аденин, называется дезоксиаденозин, производное гуанина — дезоксигуанозин. Дезоксицитидин и дезокситимидин являются дезоксинуклеозидами цитозина и тимина соответственно.
Каковы физические свойства компонентов полинуклеотидов?
В ДНК нуклеотиды присутствуют в виде 5'-фосфатных производных - dAMP, dGMP, dCMP и dTMP. Ранее указывалось, что последний нуклеотид можно обозначать просто как ТМР, так как его рибозный аналог практически не встречается.
Обе гидроксильные группы фосфорной кислоты при физиологических значения pH ионизированы, так что рКа одной ~2, а другой ~7. Это свидетельствует об их гидрофильности. Благодаря фосфатным остаткам молекула ДНК несет сильный отрицательный заряд. Сахар с его гидроксильными группами также проявляет гидрофильные свойства. Основания, наоборот, почти нерастворимы в воде и имеют ярко выраженный гидрофобный характер, хотя ряд атомов при этом сохраняет способность к образованию водородных связей.
Структура дезоксирибополинукпеотида
Мысленно отнимем от двух нуклеотидов молекулу воды. В этом случае получившаяся структура будет представлять собой динуклеотид (приведена неионизированная форма):
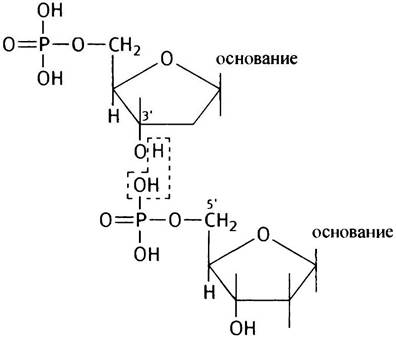
Образование динуклеотида сопровождается большим положительным изменением свободной энергии ∆G°′, следовательно, синтез не может идти путем прямой конденсации двух нуклеотидов.
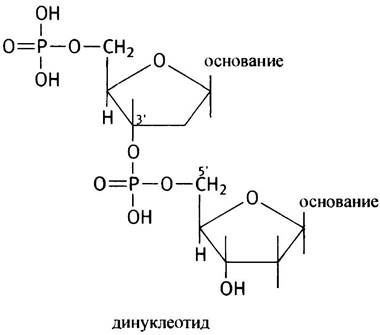
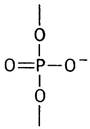
В мононуклеотиде фосфатная группа соединена с дезоксирибозой простой эфирной связью. В динуклеотиде образуется фосфодиэфирная связь. Она называется диэфирной потому, что фосфат соединен с двумя остатками углеводов:
В приведенном выше динуклеотиде это 3', 5' -фосфодиэфирная связь, где фосфат связывает 3'-ОН-группу одного нуклеотида с 5'-ОН-группой другого нуклеотида. Если далее подобным образом присоединять нуклеотиды, то получим структуру, называемую полинуклеотидом. Первичная структура ДНК - полинуклеотид огромной длины. Говоря о белках, мы отмечали наличие полипептидного остова с выступающими из него остатками аминокислот. Полинуклеотид также имеет свой остов, состоящий из чередующихся групп сахар- фосфат-сахар, а основания присоединяются к каждому остатку сахара в положении Г. Кодированная информация записана в последовательности оснований. Цепь ДНК можно схематически представить следующим образом.

Структуру цепи можно изобразить в следующем виде.
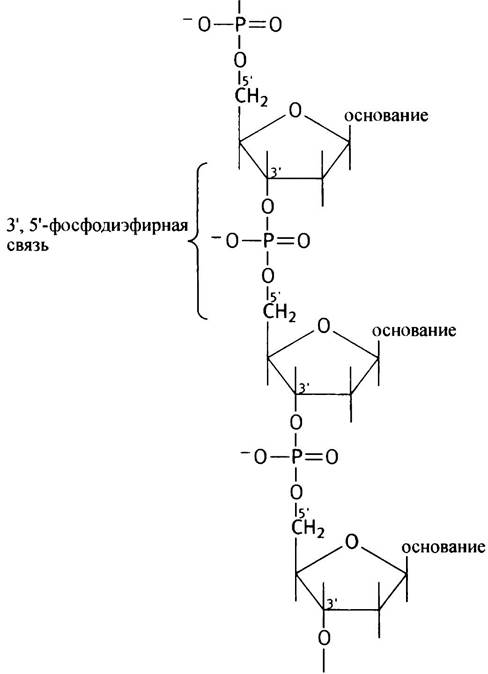
Почему дезоксирибоза?
Почему не рибоза?
В клетке присутствует ряд нуклеотидов (например, АМР), а также рибонуклеиновые кислоты, содержащие рибозу в качестве углеводного компонента. Предполагается, что рибонуклеотиды появились в ходе эволюции раньше дезоксирибонуклеотидов, а, следовательно, РНК появилась раньше ДНК. В то же время клетки идут на значительные энергетические затраты, чтобы превратить рибонуклеотиды в дезоксирибонуклеотиды, необходимые для синтеза ДНК. Почему?
Одна из весьма убедительных причин хранения генетической информации, миллионы лет накапливавшейся в виде молекул ДНК, состоит в следующем: химические молекулы всегда характеризуются определенной степенью нестабильности и способностью к спонтанному распаду. Молекулы же ДНК остаются неизменными и, в основном, интактными в течение многих поколений. Кроме того, ДНК более стабильна, чем РНК. Это связано с тем, что 2'-ОН-группа рибозы легко подвергается нуклеофильной атаке в присутствии гидроксильных ионов фосфата. В результате происходит расщепление фосфодиэфирной связи с образованием 2', 3'-циклических нуклеотидов, а в ДНК, где 2'-ОН-группы отсутствуют, этого быть не может.
Ион ОН- способствует превращению 2'-ОН-группы в группу 2'-O-, которую атакует атом фосфора. Это приводит к разрыву полинуклеотидной цепи и образованию 2',3'-циклического нуклеотида. Последующий гидролиз циклического нуклеотида дает смесь 2'- и 3'-нуклеотидов, в зависимости от места гидролиза.
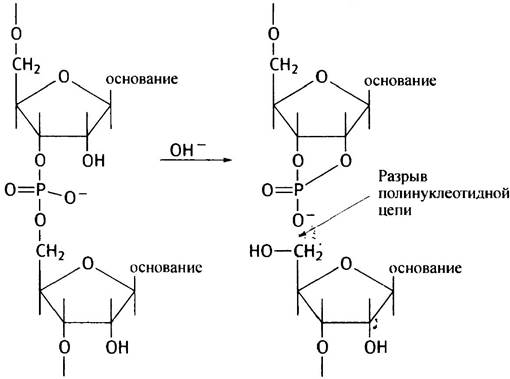
Различие в стабильности РНК и ДНК подтверждается еще и тем, что разбавленный раствор NаОН полностью разрушает РНК при комнатной температуре и не действует на ДНК. Получается, ДНК более стабильный и надежный хранитель информации, чем РНК. Заметим, что факт использования некоторыми вирусами РНК в качестве носителя информации не противоречит высказанному утверждению.
Двойная спираль ДНК
Все ДНК почти всегда существует в виде двойной цепи, за исключением ДНК некоторых вирусов. Другими словами, ДНК - это две полинуклеотидные молекулы, составляющие пару. Что же удерживает их вместе? Комплементарность пар оснований. Это означает, что, когда А и Т в двух цепях ДНК расположены друг против друга, между ними спонтанно образуются 2 водородные связи. Напомним, что водородные связи имеют малый радиус действия, поэтому для их образования требуется точное расположение пары атомов. G и С также представляют комплементарную пару, правда, между ними возникает 3 водородных связи. Другие комбинации оснований не могут быть комплементарными, поэтому G не образует пару с А или с Т и т. д. В ДНК встречаются только пары А-Т и G-С, они известны как пары оснований Уотсона-Крика. Поражает то, что образование водородных связей - полностью спонтанный процесс, не требующий ферментативного катализа. Ведь водородные
связи - слабы и легко рвутся, например, при повышении температуры. Незначительное нагревание будет сопровождаться нарушением спаривания оснований, а охлаждение - обратной ассоциацией молекул.
Геометрия пар оснований приведена на рис. 19.1.
Рис. 19.1. Водородные связи в парах оснований Уотсона- Крика
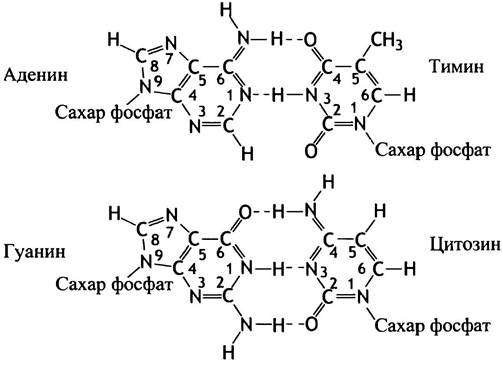
Заметим, что пара оснований всегда содержит один пурин (большая молекула) и один пиримидин (меньшая молекула); следовательно, пара оснований всегда имеет одинаковый размер. Спонтанный процесс образования пар оснований называют гибридизацией. Если молекулу ДНК разрезать на тысячи коротких двухцепочечных фрагментов длиной от 20 до 100 нуклеотидов, а затем нагреть до температуры около 95° С, спаренные цепи каждого фрагмента ДНК отделятся друг от друга: наступит плавление ДНК. Оно происходит из-за разрыва водородных связей под действием тепла и приводит к образованию одноцепочечных фрагментов ДНК с различной последовательностью оснований. Впрочем, если раствор вновь охладить, то фрагменты начнут медленно спариваться со своими прежними комплементарными цепями (рис. 19.2). Такая техника гибридизации, иногда называемая отжигом, лежит в основе молекулярной биологии гена (см. главу 24). Термодинамической движущей силой гибридизации становится высвобождение энергии, происходящее при образовании водородных связей и других слабых взаимодействиях. В наиболее стабильном состоянии величина свободной энергии минимальна. Это соответствует такому спариванию оснований, при котором появляется максимальное количество водородных связей. Экспериментальное доказательство гибридизации свидетельствует: G-С и А-Т пары оснований образуются спонтанно, и это реально существующий феномен.
Рис. 19.2. Спонтанная гибридизация комплементарных фрагментов ДНК. Для демонстрирации принципов гибридизации последовательность оснований приведена только на одном фрагменте ДНК
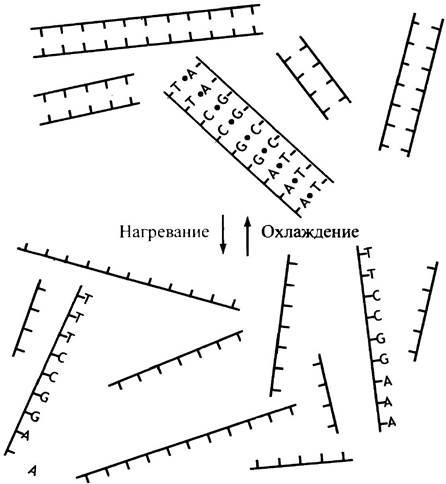
Вследствие комплементарности оснований количество G в молекуле ДНК совпадет с количеством С, а количество А - с количеством Т. Разные ДНК различаются по содержанию пар [А + Т] и [G + С] ввиду несходства их нуклеотидного состава. Это и объясняет их различия как носителей генетической информации.
В первом приближении цепь ДНК можно представить в виде длинных сплошных линий, соответствующих сахарофосфатному остову; удерживаемые водородными связями основания всегда ориентированы внутрь.
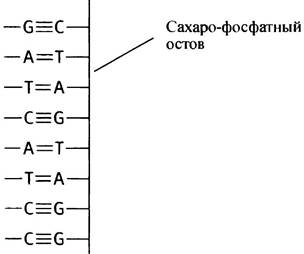
Заметим, что на участках ДНК, богатых [G + С], две цепи будут удерживаться вместе сильнее, чем на участке, где больше пар [А + Т].
Однако слабые взаимодействия оказывают существенное влияние на конформацию крупных молекул, и ДНК здесь - не исключение. Поэтому приведенная выше структура с прямыми цепями в нормальных условиях не встречается.
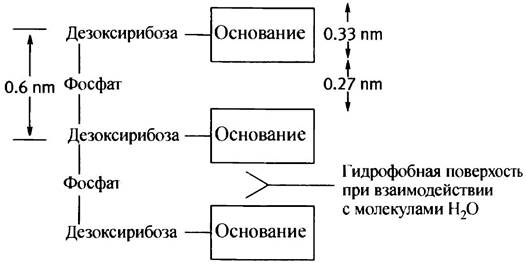
Длина фосфодиэфирной связи составляет 0,6 нм (1 нанометр = 10-9 м), а толщина оснований - около 0,33 нм, так что в прямой, похожей на лестницу структуре между ними должны оставаться промежутки. Плоскости оснований, как уже говорилось, гидрофобны. В «прямой» структуре они вынуждены находиться в водном окружении, что придает молекуле нестабильность. Что же можно сделать для связи между собой гидрофобных оснований и исключения их контакта с Н2O? Нужно уложить их друг на друга, изогнув при этом фосфодиэфирную связь; тогда гидрофобные взаимодействия между плоскостями оснований будут способствовать их укладке (рис. 19.3, а).
Каждая из лежащих в одной плоскости пар оснований параллельна соседним близлежащим парам, и все они образуют так называемую стопку оснований. Причем основания ориентированы таким образом, что между противоположными цепями могут образоваться водородные связи. Впрочем, такая «лестничная» структура не встречается в ДНК, поскольку невозможна стереохимически. На самом деле две цепи ДНК образуют спираль, в которой основания расположены внутри, а гидрофильные сахарные и фосфатные группы снаружи (рис. 19.3, б, в).
Рис. 19.3. Модель укладки нуклеотидов в ДНК: а - Тип связи пар оснований в структуре «наклонной лестницы»; б - контур остова в двойной спирали ДНК; в - то же, что и б, изображены пары оснований в центре спирали (каждый цилиндр - пара оснований); г - расположение двух соседних пар оснований в двойной спирали. Показано смещение, определяемое структурой двойной спирали. S - сахар, Р - фосфат, В - основание. На рис. 19.4 приведена более подробная модель, соответствующая в
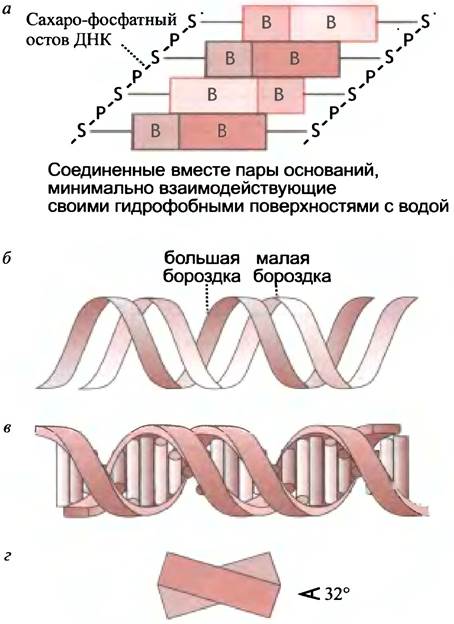
Благодаря такой структуре обеспечивается тесное взаимодействие пар оснований и исключается их контакт с водой, но стопка оснований уже не абсолютно вертикальна. Пары оснований слегка смещены относительно друг друга; как показано на рис. 19.3, г, на один виток приходится около 10 пар.
Спираль правозакрученная; если смотреть вдоль ее оси, то повороты следуют по часовой стрелке (представьте, что вы закручиваете винт, держа отвертку в правой руке: направление вращения дает представление о направлении поворота спирали). Структура двойной спирали
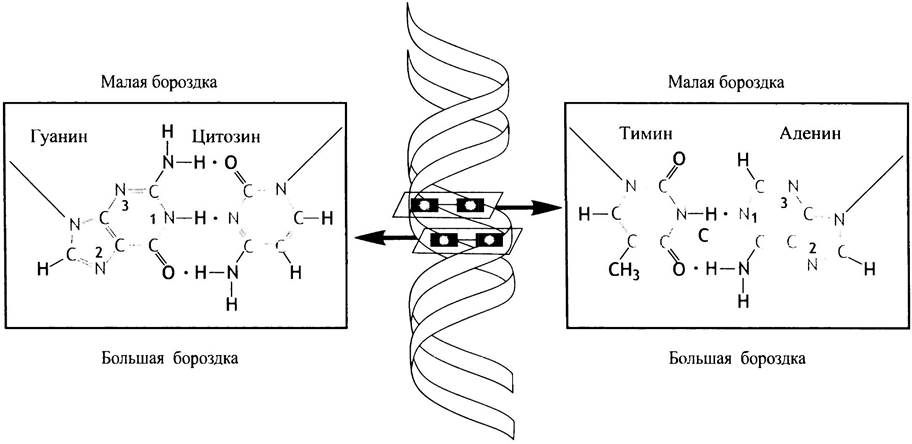
такова, что она имеет две бороздки - большую и малую (см. рис. 19.3, б). Хотя бороздки показаны на плоскости, их трудно изобразить вне трехмерного пространства. Если у вас есть возможность ознакомиться с объемной моделью ДНК, это поможет понять направление спирали и обнаружить на ней две бороздки, что чрезвычайно важно. Часть каждого основания видна как из большой, так и из малой бороздок: это делает их доступными для взаимодействия с другими молекулами. Большая бороздка обеспечивает более легкое взаимодействие с белками, которые «узнают» соответствующие основания. Пары оснований совершенно не похожи, если их рассматривать со стороны двух разных бороздок (рис. 19.4); значение этого факта будет разъяснено в главе 21, где обсуждается генетическая регуляция.
Рис. 19.4. Схематичное изображение, показывающее, что со стороны большой и малой бороздок одна и та же пара оснований в ДНК выглядит по-разному
ДНК-связывающие белки, предназначенные для узнавания специфических последовательностей в ДНК, могут присоединяться к соответствующим участкам без раскручивания спирали
Описанная выше конформация ДНК известна как В-форма (рис. 19.5): именно в такой форме ДНК обычно находится в клетке. Однако ДНК может изменять свою конфигурацию в зависимости от условий. При дегидратации двойная спираль приобретает более сплющенную форму с большим углом наклона оснований, и в этом случае ее называют A-формой. В такой форме ДНК встречается в спорах. Известна и другая конфигурация - Z-форма, когда сахарофосфатный остов образует зигзагообразную (англ. zigzag) линию вдоль спирали. В Z-форме двойная спираль закручена влево, а в В-форме - вправо. Есть сведения, что Z-форма встречается в условиях высокой ионной силы раствора в коротких молекулах синтетической ДНК с чередующимися попеременно пуриновыми и пиримидиновыми основаниями. Биологическое значение А- и Z-формы пока неизвестно, однако не исключена возможность приспособительного изменения конфигурации ДНК на коротких участках хромосом.
Рис. 19.5. Модель В-формы ДНК. Представлена трехмерная модель фрагмента ДНК с двумя большими и одной малой бороздками
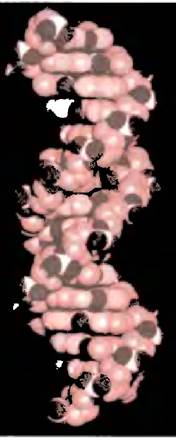
Важным свойством двойной спирали является ее способность изгибаться. Молекула ДНК может быть в миллион раз длиннее, чем клетки или ядра самых больших размеров; чтобы поместиться в них, молекула должна быть гибкой.
Следует отметить, что, хотя ДНК почти всегда находится в форме двойной спирали, существуют и одноцепочечные ДНК, например, в некоторых вирусах бактерий. Тогда для обеспечения термодинамической стабильности молекулы образуется сложная скрученная структура. Главное, что и в этом случае жизненный цикл вируса включает стадию, когда ДНК находится в форме двойной спирали (см. главу 23); следовательно, основные принципы передачи генетической информации, предусматривающие образование комплементарных пар оснований, остаются в силе.
Цепи ДНК антипараллельны: что это значит?
Под антипараллельной мы понимаем структуру, в которой две цепи в двойной спирали ДНК имеют противоположную направленность. Две антипараллельные цепи ДНК приведены на рис. 19.6.
Рис. 19.6. Две антипараллельные цепи ДНК. В - основания
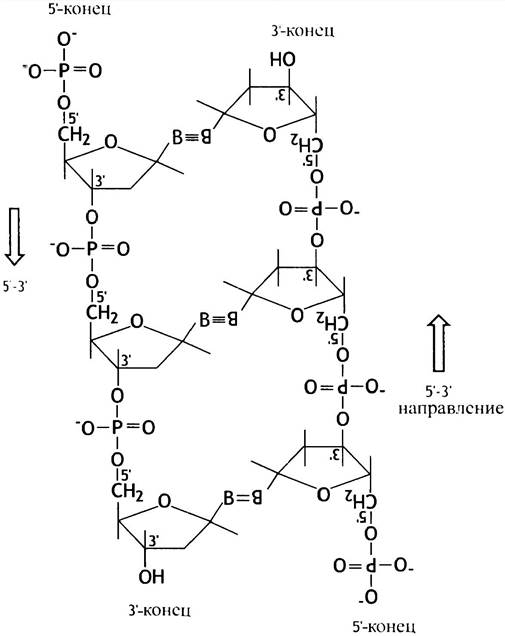
Что же означает полярность, или направление цепи ДНК? Каждая линейная цепь имеет два конца, на одном из них расположена 5-ОН-группа дезоксирибозы (не связаная с другим нуклеотидом). Иногда она бывает фосфорилирована. Это - 5'-конец. На другом расположена свободная 3'-ОН-группа (она также не связана с другим нуклеотидом и может быть этерифицирована фосфатной группой). Это - 3'-конец. Поэтому на каждом конце двойной спирали всегда есть один 5'-конец и один 3'-конец. Вы никогда не увидите два 5'-конца или два 3'-конца вместе! Это и означает антипараллельность. Если фрагмент ДНК имеет кольцевую форму (свободные концы отсутствуют), полярность отдельных нитей все равно сохраняется. Это можно проиллюстрировать на примере расположения в цепях углеводных компонентов.
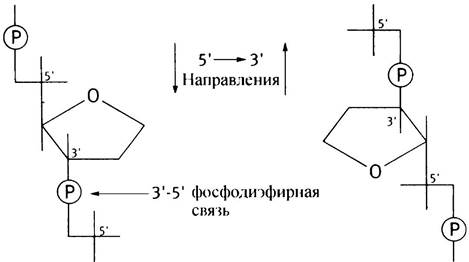
Вы можете узнать направление, ориентируясь на положения 5' и 3'. Так, на приведенной слева структуре цепь идет в направлении 5’ —> 3' сверху вниз, а справа - 5' —> 3' соответствует направлению снизувверх (условимся обозначать направление последовательности нуклеотидов в ДНК от 5' —> 3').
Обратите внимание на кажущееся противоречие! Когда вы движетесь по цепи ДНК в направлении 5' —> 3' (от 5'-конца к 3’-концу), каждая фосфодиэфирная связь, которую вы «проходите», образована между 3'-ОН-группой дезоксирибозы предыдущего и 5'-ОН-груп- пой дезоксирибозы последующего нуклеотида. Это означает, что - исходя из положения фосфата - вы следуете от 3'-конца к 5'-концу цепи. Следует помнить, что направление 5' —> 3' для нити линейной молекулы ДНК означает, что вы двигаетесь от концевой 5'-ОН-группы к концевой 3'-ОН-группе, или от 5'-группы одного остатка дезоксирибозы к 3'-группе соседнего. Это необходимо запомнить, как в силу важности самого вопроса, так и для понимания материала последующих глав.
Антипараллельное расположение цепей предъявляет определенные стереохимические требования к структуре двойной спирали.
Существуют правила написания последовательности расположения нуклеотидов в ДНК. Простую полинуклеотидную структуру принято изображать в виде последовательности букв, обозначающих соответствующие основания нуклеотидов. А иногда в виде буквы «р», располагаемой между основаниями, указывается фосфодиэфирная связь (например, СрАрТрGр, и т. д.).
Предположим, у нас имеется фрагмент двухцепочечной ДНК со следующей последовательностью оснований:
5' САТGТА 3'
3' GТАСАТ 5'
Иногда записывают последовательность для каждой цепи, но в большинстве случаев нет необходимости изображать обе цепи, так как вторая комплементарна
первой. Поэтому, несмотря на наличие в гене двух цепей, его структуру часто представляют в виде простой последовательности оснований. При условии, что она записывается с 5'-конца слева, специального обозначения 5'- и 3'-концов не требуется. Таким образом, приведенная выше структура может быть обозначена как CATGTA.
Каковы размеры молекулы ДНК?
Размеры молекул ДНК, если ориентироваться по числу входящих в их состав оснований, сильно различаются. Но даже самые короткие из них представляют собой очень крупные молекулы, а молекулы ДНК с наиболее длинными цепями просто гигантские. Небольшие ДНК вирусов состоят из нескольких тысяч пар оснований; ДНК Е. coli содержит около 4 млн пар оснований. Геном человека (полная сумма ДНК хромосом) достигает длины 1-2 м и включает 6 млрд пар оснований. Если учесть, что человек состоит из 1013клеток, и подсчитать суммарную длину его ДНК, получится астрономическая величина, сравнимая с диаметром солнечной системы.
Ясно, что чем больше информации необходимо сохранить, тем большее количество ДНК требуется. Следовательно, чем сложнее организм, тем большее количество ДНК содержат его клетки. Впрочем, это соблюдается не всегда. Например, крупные клетки бобов содержат больше ДНК, чем клетки человека, а у земноводных ее больше, чем у других позвоночных. Появление аномалий (ошибок) в структуре ДНК приводит к потере некоторых функциональных свойств.
Каким образом ДНК упакована в ядре?
В биохимии часто приходится сталкиваться с поразительными решениями эволюцией тех или иных проблем! Упаковка ДНК — одна из них.
В клетках человека содержится 46 хромосом, а общая длина заключенной в клетке ДНК составляет 1-2 м, причем она упакована в ядре, диаметр которого в миллионы раз меньше. Значит, требуется тщательно продуманная система укладки. В эукариотических клетках ДНК находится в составе хроматина - комплекса ДНК с белками. Белки представлены преимущественно гистонами - основными белками, богатыми аргинином и лизином: благодаря своему положительному заряду они образуют ионные связи с расположенными на внешней стороне двойной спирали ДНК отрицательно заряженными фосфатными группами. Аминокислотная последовательность эукариотических гистонов высоко консервативна. Например, первичная структура гистонов, Выделенных из растения (горох) и млекопитающего (корова), отличается только на две аминокислоты, да и эти замены не очень существенны (валин на изолейцин, лизин на аргинин). Столь выраженное постоянство структуры позволяет предположить, что такой белок должен точно взаимодействовать с настолько же консервативной структурой или структурами.
Начнем с рассмотрения четырех гистонов - Н2А, Н2В, НЗ и Н4. Они образуют октамерный белковый комплекс, называемый нуклеосомной сердцевиной (англ. nucléosome соге), вокруг которого молекула ДНК совершает два оборота (около 146 пар оснований). Такой комплекс октамера гистоновых белков с ДНК является основной структурной единицей хроматина и называется нуклеосомой. Отдельные нуклеосомы соединяются линейными участками ДНК - линкерами (рис. 19.7, а, б). Если хроматин в пробирке обработать ферментом, выделенным из микроорганизмов и способным Гидролизовать ДНК на нуклеотиды, линейные участки ДНК действительно подвергаются гидролизу, а около 146 пар оснований, «намотанных» на нуклеосомную сердцевину, оказываются защищенными от действия фермента (он не в состоянии гидролизовать эту часть ДНК). Экстрагированная после такой обработки ДНК содержит неповрежденные фрагменты. Это наблюдение легло в основу представлений о нуклеосомной структуре.
Нуклеосомы имеют примерно 10 нм в диаметре и образуют нити толщиной 10 нм (см. рис. 19.7, б). И все же такой упаковки ДНК недостаточно, в eе дальнейшей организации участвует пятый гистон: он не входит в состав нуклеосомы и роль его не совсем ясна. Далее нить 10 нм толщиной конденсируется с образованием нити толщиной 30 нм (рис. 19.7, в). Такая упаковка нуклеосом зачастую описывается как соленоидная; впрочем, это лишь одна из моделей. Согласно другой модели, упаковка имеет зигзагообразную форму. Поскольку точная структура соленоида еще не установлена, схема 30-нанометровой нити приводится без деталей упаковки. Электронная микрофотография такой нити представлена на рис. 19.8. При этом ДНК конденсируется в 100 раз, т. е. происходит стократное уменьшение ее длины. Нити толщиной 30 нм образуют длинные петли, прикрепленные к центральному поддерживающему белку хромосом (рис 19.7, г). За счет формирования петель структура становится еще более компактной, но пока не выяснено, включает ли такое скручивание дополнительную спирализацию, обеспечивающую 10 000-кратную конденсацию исходной молекулы ДНК.
Рис. 19.7 Модель упаковки хроматина у эукариот: а - Нуклеосома; б - нуклеосомы с линкерными участками ДНК; в - нить хроматина 30 нм толщиной; г - петли этой нити, прикрепленные к центральному поддерживающему белку. Петли далее конденсируются путем сверх- спирализации, образуя чрезвычайно компактную метафазную хромосому. Последняя стадия конденсации на рисунке не представлена
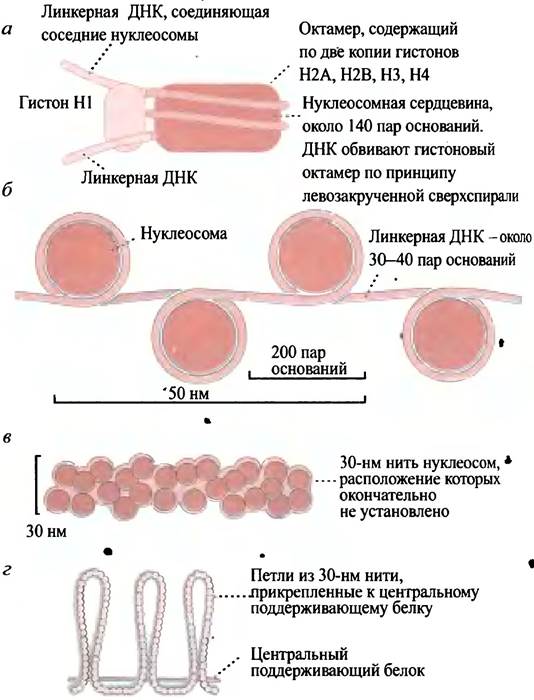
Рис. 19.8. Электронная микрофотография нити хроматина 30 нм толщины
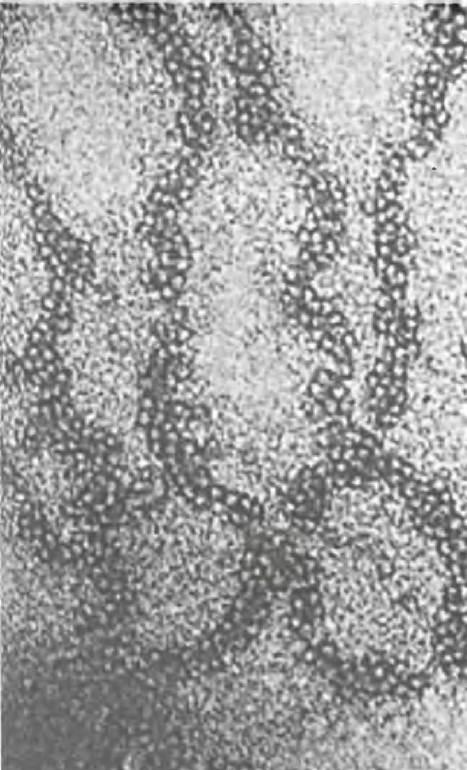
Таким образом, есть несколько уровней организации ДНК: 1) закручивание вокруг нуклеосом; 2) упаковка нуклеосом в 30-нанометровую нить, объединяющую 3-5 нуклеосом; 3) образование петли длиной в тысячи нуклеосом, прикрепленной к центральной поддерживающей структуре; 4) формирование из петли спирали и/или складки (см. рис. 19.7, а, г).
Для выполнения своей функции упакованная ДНК должна быть доступна ферментам, участвующим в считывании генетической информации и репликации.
В клетках Е. coli двойная спираль ДНК имеет кольцевую форму. Она длиннее клетки примерно в 1000 раз. У прокариот нет ядерной мембраны и нет упорядоченных структур наподобие эукариотических нуклеосом. Тем не менее, в Е. coli найдены гистоноподобные белки, которые, предположительно, выполняют сходную функцию. Поскольку в бактериальных клетках белки связаны с ДНК менее прочно, чем эукариотические гистоны, нуклеосомоподобные комплексы в них не обнаруживаются. Петельная структура ДНК в Е. coli стабилизируется белками, но детали этого процесса до конца не ясны.
Как структура ДНК соотносится с компактными хромосомами эукариот, видимыми в световой микроскоп?
Хромосомы в окрашенных делящихся эукариотических клетках видны в световом микроскопе как компактные плотные образования, подобные тем, что изображены на рис. 19.9.
Рис. 19.9. Хромосома в метафазе состоит из двух дочерних хроматид
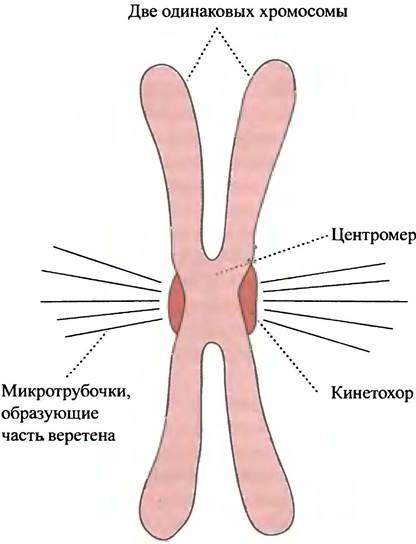
Компактные структуры в делящихся клетках представляют так называемые метафазные хромосомы. Там ДНК находится в наиболее конденсированном состоянии, что помогает перемещению хромосом после их деления в каждую из дочерних клеток.
Наиболее плотно упакованная область ДНК, состоящая из повторяющихся последовательностей, приходится на центромеру. Она удерживает вместе пару хроматид, а на их внешних поверхностях располагаются комплексы белков кинетохора. Для разделения дочерних хромосом к кинетохору прикрепляются микротрубочки митотического веретена (см. главу 29). После деления клетки хромосомы становятся менее упакованными; их называют интерфазными хромосомами. ДНК метафазных хромосом функционально инертна, а интерфазных функционально активна (по сравнению с простой нитью ДНК интерфазные хромосомы более упакованы). Степень конденсации ДНК в эукариотических клетках очень важна. Прежде чем использовать содержащуюся в молекулах информацию, ДНК должна стать доступна ферментам и другим белкам и не иметь плотно упакованной структуры. Определенные участки эукариотических хромосом содержат ДНК, которая, предположительно, не входит в структуру гена, т. е. не несет информации, необходимой для синтеза белка. ДНК в этих местах даже в интерфазных хромосомах остается конденсированной в виде так называемого гетерохроматина, в то время как функционирующие области - эухроматин - менее конденсированы. Функция гетерохроматина до конца не ясна.
В хроматине помимо гистонов присутствуют и другие, негистоновые белки. Они участвуют в образовании длинных петель ДНК, прикрепленных к осевым поддерживающим белковым структурам. Таким образом, хроматин является сложной структурой. В зависимости от экспрессии гена меняются и степень его упаковки, и степень упаковки самой ДНК в хроматине. Термин экспрессия гена относится к процессу синтеза белка, контролируемого геном, и также используется в случаях кодирования генами специфических молекул РНК (см. с. 283).
Что такое ген в молекулярных терминах?
Ген - единица наследственности. Он представляет собой часть гигантской молекулы ДНК и содержит закодированную информацию о последовательности аминокислот одной полипептидной цепи (см. с. 264, где дается детальное описание гена). Информация хранится в виде последовательности оснований ДНК, и один ген отличается от другого только закодированной информацией. Для большей наглядности можно провести аналогию между хромосомой и магнитной лентой: тогда ген сравним с фрагментом музыкальной записи, а последовательность оснований - с магнитными сигналами. Выше сказанное относится к «стандартным» (типичным) кодирующим белок генам. Их подавляющее большинство.
Некоторые вариации «стандартного» гена
Иногда существуют множественные копии одного и того же гена. Например, гены некоторых гистоновых белков: сотни тандемно повторяющихся копий обнаружены у морских ежей. Это может быть результатом дупликации гена в процессе эволюции. Множественные копии гена способны обеспечивать ускоренный синтез белка. В эволюционном плане это позволяет приспособить одну из копий к выполнению какой-нибудь иной (альтернативной) функции. Существуют также псевдогены - копии функциональных генов, способные к экспрессии, синтезирующие нефункциональный белок или белковый фрагмент.
Есть группа генов, которые не кодируют белков: это гены рибосомной и транспортной РНК. Они кодируют молекулы РНК, необходимые для синтеза белков (см. главу 22).
Обычный «стандартный» ген представляет собой фрагмент ДНК, расположенный в определенном месте хромосомы. Последовательности в ДНК не всегда жестко фиксированы; иногда в них имеются фрагменты, способные перемещаться из одного места ДНК в другое. Они называются подвижными, или прыгающими генами (см. с. 259). Хромосомы также могут включать новые гены из ретровирусов (см. с. 313).
Структура гена не всегда постоянна. При определенных условиях отрезок ДНК, соответствующий какому-либо гену, может амплифицироваться, так что получится огромное количество копий гена и соответственно резко увеличится (примерно в 1000 раз) синтез закодированного в нем белка. Такое явление наблюдается в экспериментах на культуре клеток рака мышей, обработанных метотрексатом - лекарственным препаратом «антифолатного» действия, используемым для лечения лейкемий (см. с. 226).
Повторяющаяся ДНК
В клетках человека не вся ДНК находится в виде генов. Например, в наших хромосомах существуют довольно протяженные (длиной в несколько сотен оснований) участки ДНК, повторяющиеся сотни и тысячи раз, разбросанные по всей длине хромосомы. Примером могут служить Alu-последовательности (Alu - аббревиатура полного названия фермента, осуществляющего их гидролиз). Функция Alu-последовательностей не установлена.
Еще один вид повторяющихся последовательностей ДНК - сателлитная ДНК: она часто встречается в области центромеры хромосом и является частью гетерохроматина. Термин сателлитная возник потому, что гигантская молекула эукариотической ДНК при выделении обычным способом расщеплялась на фрагменты. При их анализе в градиенте плотности большая часть ДНК оседала одной полосой, в то время как повторяющиеся участки ДНК, имеющие другую плотность, отделялись от предыдущей полосы (т. е. выявлялись как сателлиты). Сателлитная ДНК состоит из коротких последовательностей, повторяющихся в тандемном порядке (друг за другом). Длина каждого фрагмента примерно 105 оснований. Другие повторяющиеся последовательности ДНК рассеяны по хромосоме.
Где мы сейчас находимся?
Дабы не потерять основной нити рассуждения среди обилия деталей, напомним, что мы сначала имели дело со структурой белка, после чего перешли к вопросам, связанным с процессом его биосинтеза. Теперь мы обратимся к биосинтезу ДНК. Во-первых, это логичное продолжение изучения структуры ДНК; во-вторых, поможет пониманию последующих этапов синтеза белка. Поэтому в следующей главе мы обсудим синтез ДНК, а после перейдем к изучению путей передачи генетической информации, непосредственно связанной с биосинтезом белка.
Вопросы к главе 19
1. Напишите структуру динуклеотида.
2. Рибонуклеиновая кислота (РНК) появилась раньше дезоксирибонуклеиновой кислоты. Почему появилась ДНК?
3. Основания в ДНК гидрофобны. Объясните, как это влияет на структуру двуцепочечной ДНК.
4. Назовите основные характеристики двойной спирали ДНК. Это лево- или правозакрученная спираль? Сколько пар оснований формируют один виток спирали ДНК?
5. Объясните, что означает антипараллельность цепей ДНК в двойной спирали.
6. Объясните, что означает направление 5' —> 3' в линейной молекуле ДНК.
7. Если вы видите структуру ДНК, записанную как САТАGССG, что она означает, исходя из структуры двойной спирали и полярности ее цепей? Объясните свой ответ.
8. Что такое нуклеосома? Что подтверждает ее существование?