Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Стехиометрия и энергетика метаболических превращений
Стехиометрия клеточного роста и образования продуктов метаболизма
Общая стехиометрия клеточного роста; состав среды и коэффициенты выхода
В процессе роста клетки потребляют субстраты, которые обеспечивают их энергией и исходными веществами для синтеза новой клеточной массы. С самой общей точки зрения для реализации этого процесса необходимо, чтобы в питательной среде находились все химические элементы, требующиеся для формирования новой клеточной массы, и чтобы свободная энергия усвоенных субстратов превышала свободную энергию образующихся клеток и продуктов метаболизма (рис. 5.28)*. Другими словами, свободная энергия образующихся веществ должна быть меньше свободной энергии утилизированных субстратов.
Кроме того, питательные вещества должны иметь соответствующее молекулярное строение, определяемое специфичностью клеточных ферментов, участвующих в процессах катаболизма и биосинтеза. В то же время при изучении роста клеток как единого процесса (а именно с этой точки зрения мы будем изучать его в большей части настоящего раздела) привлечение деталей механизмов метаболических превращений может оказаться не только ненужным, но и вредным. Основной смысл изображенного на рис. 5.28 процесса роста клеток как единого явления заключается в том, что ему свойственны определенные стехиометрические ограничения независимо от того, какой конкретный механизм или какие метаболические пути использует «система» клеток для достижения цели «реакции» — роста клеток. Поэтому для решения вопроса о пригодности тех или иных питательных веществ мы просто будем рассматривать только те питательные вещества, которые заведомо являются подходящими субстратами для роста изучаемого штамма клеток.
* В настоящем разделе продуктами метаболизма мы будем называть органические вещества, отличающиеся от продуктов жизнедеятельности клеток, выделяемых ими в среду. Под питательными веществами или субстратами мы будем подразумевать вещества, потребляемые клеткой из среды в процессе роста или образования продуктов метаболизма.
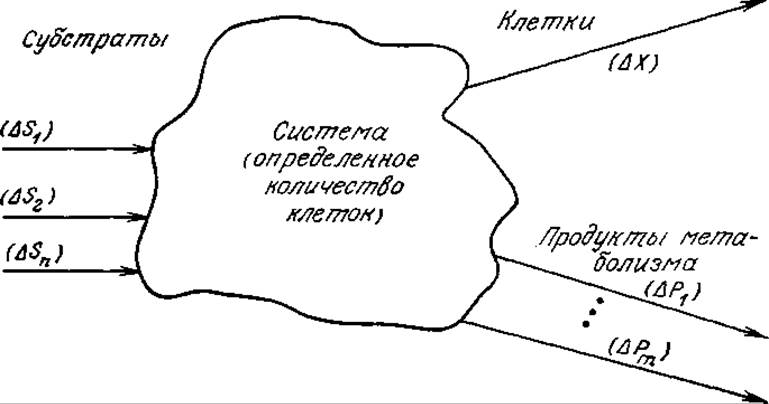
РИС. 5.28. Клеточный рост как единый процесс. Под «системой» здесь подразумевается определенное количество клеток. В процессе роста «система» выполняет функции катализатора, превращая субстраты в новые клетки и продукты метаболизма.
Простейшая форма стехиометрических ограничений, накладываемых на состав питательной среды, в краткой форме может быть выражена следующим образом: если мы хотим вырастить общую массу клеток (X) (прописными буквами в скобках мы будем обозначать общую массу компонента, указанного в скобках) и если весовая доля элемента і в клетке равна то минимально необходимое общее содержание элемента і во всех субстратах должно составлять wi(X). В общем случае подобные вычисления лучше выполнять в расчете на массу сухого клеточного вещества. При выполнении таких расчетов для различных элементов необходимо знать их относительное содержание в клетке. Примером могут служить довольно подробные данные об элементном составе бактерии Е. coli, приведенные в табл. 2.1, а в табл. 5.10 указано содержание основных элементов (С, N, О и Н) в клетках целого ряда микроорганизмов. Хотя элементный состав клеток несколько изменяется в зависимости от вида микроорганизма и даже от состава питательной среды з случае одного и того же штамма, в целом эти данные вполне надежно определяют элементный состав клеточной массы. Смысл приведенных в этой таблице эмпирических формул клеток и их применение мы рассмотрим в следующем разделе.
Таблица 5.10. Элементный состав некоторых микроорганизмов. Символом μ, обозначена удельная скорость роста, равная массе клеток, образующихся из данной массы клеток в единицу времени (см. гл. 7)а
|
Микроорганизм |
Лимитирующее питательное вещество |
Состав, масс. % |
|||||||||
μ ч-1 |
С |
H |
N |
O |
P |
S |
Зола |
Эмпирическая химическая формула |
Молекулярная» масса, отвечающая эмпирической формуле |
||
Бактерия |
53,0 |
7,3 |
12,0 |
19,0 |
8 |
СН1,666N0,20O0,27 |
20,7 |
||||
Бактерия |
47,1 |
7,8 |
13,7 |
31,3 |
СН2N0,25О0,5 |
25,5 |
|||||
Aerobacter aerogenes |
48,7 |
7,3 |
13,9 |
21,1 |
8,9 |
CH1,78N0,24O0,33 |
22,5 |
||||
Klebsiella aerogenes |
Глицерин |
0,1 |
50,6 |
7,3 |
13,0 |
29,0 |
CH1,74N0,22O0,43 |
23,7 |
|||
К. aerogenes |
Глицерин |
0,85 |
50,1 |
7,3 |
14,0 |
28,7 |
CH1,73N0,24O0,43 |
24,0 |
|||
Дрожжи |
47,0 |
6,5 |
7,5 |
31,0 |
8 |
СН1,66N0,13О0,40 |
23,5 |
||||
Дрожжи |
50,3 |
7,4 |
8,8 |
33,5 |
CH1,75N0,15O0,5 |
23,9 |
|||||
Дрожжи |
44,7 |
6,2 |
8,5 |
31,2 |
1,08 |
0,6 |
CH1,64N0,16O0,52P0,01S0,005 |
26,9 |
|||
Candida utilis |
Глюкоза |
0,08 |
50,0 |
7,6 |
11,1 |
31,3 |
CH1,82N0,19O0,47 |
24,0 |
|||
C. utilis |
Глюкоза |
0,45 |
46,9 |
7,2 |
10,9 |
35,0 |
CH1,84N0,2О0,56 |
25,6 |
|||
C. utilis |
Этанол |
0,06 |
50,3 |
7,7 |
11,0 |
30,8 |
CH1,82N0,19O0,46 |
23,9 |
|||
C. utilis |
Этанол |
0,43 |
47,2 |
7,3 |
11,0 |
34,6 |
CH1,84N0,2O0,56 |
25,5 |
|||
* Воспроизведено с разрешения из работы: Atkinson В., Mavituna F., Biochemical Engineering and Biotechnology Handbook, p. 120, Macmillan Publishers Ltd., Surrey, England, 1983.
На самом деле определить необходимый для эффективного роста клеток состав среды значительно сложнее, поскольку изложенный выше основной стехиометрический принцип не учитывает, во-первых, то, что часть химических элементов субстратов входит в выделяемые в среду продукты метаболизма, во-вторых, ограничений, накладываемых скоростями реакций, в-третьих, особых метаболических свойств данного штамма клеток, обусловленных ингибирующими эффектами каких-либо питательных веществ или продуктов метаболизма. Первую причину осложнений мы рассмотрим позднее в этом же разделе, а вторую обсудим сейчас. Затем мы перейдем к изучению экономических коэффициентов — простейших параметров, с помощью которых можно количественно оценить стехиометрию роста клеток.
Хотя в этом разделе основное внимание мы постараемся уделять именно стехиометрии, при разработке подходов к упрощению стехиометрических выражений клеточного роста и образования продуктов метаболизма нам придется принимать во внимание и кинетические факторы. Число питательных веществ в среде обычно очень велико, и учесть все эти вещества в анализе стехиометрии или кинетики процесса практически невозможно. Поэтому обычно стараются описать систему упрощенно, на базе ограниченного числа лимитирующих процесс ключевых соединений или элементов. Найти стехиометрически лимитирующее соединение можно на основе стехиометрии клеточного роста. Для этого необходимо определить, какой субстрат первым полностью израсходуется в «реакции» клеточного роста, изображенной на рис. 5.28. Лимитирующие соединения других типов можно идентифицировать путем изучения влияния состава среды на рост клеток. Для данного штамма в данных условиях (температура, pH, присутствие других питательных веществ) лимитирующий скорость роста компонент питательной среды можно найти экспериментально следующим образом. Предположим, что клетки растут в определенной среде в заданных условиях. В этой системе резко повышают концентрацию одного из компонентов среды и затем определяют изменение скорости роста клеток. Часто состав среды подбирают так, чтобы скорость клеточного роста лимитировал только один компонент, а относительно небольшие (по сравнению с базовыми значениями) изменения концентраций всех других компонентов среды практически не влияли бы на скорость роста клеток. В принципе, однако, возможны и такие варианты, когда скорость клеточного роста определяют несколько субстратов одновременно.
Компонент, лимитирующий скорость клеточного роста, и стехиометрически лимитирующий компонент не обязательно должны быть одним и тем же веществом. Другими словами, в определенных условиях скорость роста клеток может ограничивать одно соединение, но рост клеток в периодическом процессе может прекратиться в силу истощения запасов другого соединения. В то же время в различных математических выражениях, описывающих рост микроорганизмов количественно, такая возможность часто игнорируется, что может приводить к известным трудностям при изучении режима биореакторов и неправильным выводам при интерпретации их характеристик. Не забывая о возможных осложнениях такого рода, мы тем не менее будем считать, что клеточный рост лимитирует только одно питательное вещество. Если речь идет о конкретной системе и определенной среде, то всегда можно экспериментально проверить справедливость допущения о том, что лимитирующий скорость роста и стехиометрически лимитирующий компоненты являются одним и тем же веществом.
Часто оказывается, что общее количество клеточной массы, образующейся в процессе клеточного роста, пропорционально массе утилизированного клетками субстрата (обычно углеродсодержащего предшественника, источника энергии или кислорода) (рис. 5.29). Экономическим коэффициентом называют соответствующее соотношение:
![]()
Следует отметить, что размерность экономического коэффициента определяется способом выражения количества клеточной массы и количества субстрата (например, в единицах массы или в числе молей); этот факт всегда следует учитывать при использовании опубликованных в литературе значений У. При одновременной утилизации из среды нескольких различных субстратов Si можно определить несколько соответствующих экономических коэффициентов клеточного роста Yх/si. В табл. 5.11 приведены экономические коэффициенты для роста ряда микроорганизмов в средах, отличающихся природой источника углерода; здесь цифры в одной строке отвечают разным способам измерения количества утилизированного субстрата (в граммах, молях или в граммах углерода в 1 моль субстрата).
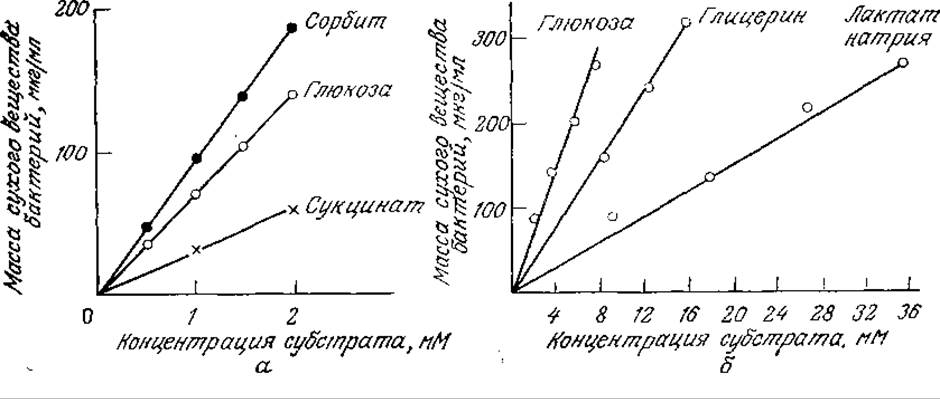
РИС. 5.29. Зависимость общего количества образующейся биомассы от количества утилизированного субстрата в случае роста бактерий Aerobacter aero- genes в аэробных условиях (а) и бактерий Propionibacterium pentosaceum в анаэробных условиях (б) на различных источниках углерода. [Воспроизведено с разрешения из работ: a) Hadjipetrou L. Р., Gerrits J. Р., Tenlings F. A. G., Stouthamer А. H., Relation between Energy Production and Growth of Aerobacter aerogenes, J. Gen. Microbiol., 36, 139 (1964); 6) Bauchop T., Elsden S. R., Growth of Microorganisms in Relation to their Energy Supply, J. Gen. Microbiol., 23, 457 (1960).]
Если для данной культуры клеток в данной питательной среде экономический коэффициент приблизительно постоянен, то уравнение (5.46) позволяет определить изменение концентрации клеточной массы по изменению концентрации субстрата, и наоборот. Иными словами, одну из этих переменных можно выразить через другую и при расчетах в ходе проектирования реакторов одну переменную можно не учитывать. Это стехиометрическое соотношение может пригодиться и при контроле работы биологических реакторов; в данном случае концентрация субстрата может служить одновременно и мерой концентрации клеточной массы, если только известны обе соответствующие исходные величины. Это чрезвычайно удобно, поскольку обычно концентрации субстратов определять значительно легче, чем концентрацию биомассы. На практике удобнее всего определять концентрации веществ, покидающих зону аэробного биореактора, например СO2. В этом случае в качестве основы для установления зависимостей между различными переменными биотехнологических процессов также используются стехиометрические соотношения. Однако, прежде чем обратиться к изучению таких зависимостей, необходимо упомянуть о вариабельности экономического коэффициента и о понятии метаболизма поддержания.
Нет никакой гарантии, что найденный эмпирическим путем экономический коэффициент, представляющий собой кажущееся стехиометрическое соотношение, всегда постоянен для данного организма в данной среде. Действительно, экспериментально установлено, что часто экономический коэффициент изменяется при изменении скорости роста клеток. Чтобы понять причину этих изменений, иногда полезно разбить процесс утилизации субстрата на три части: превращение субстрата в клеточную массу (ассимиляция), обеспечение энергии для клеточного синтеза и обеспечение энергии для поддержания культуры. Под энергией поддержания здесь подразумеваются энергетические затраты клетки, необходимые для ее выживания или сохранения в определенном (потенциально) жизнеспособном состоянии, но не обеспечивающие клеточный рост и образование новых клеток. Эта энергия расходуется на активный транспорт ионов и других веществ через клеточные мембраны и на возобновление деградирующих составных частей клетки.
Таблица 5.11. Экономические коэффициенты для аэробного роста ряда микроорганизмов на различных источниках углерода. Здесь символом YX/O2 обозначен коэффициент, выраженный в граммах клеточной массы в расчете на 1 г усвоенного O2а
|
YX/S |
|||||
Микроорганизм |
Субстрат |
г/г |
г/моль |
г/г (углерода) |
YX/O2, г/г |
|
Aerobacter aerogenes |
Мальтоза |
0,46 |
149,2 |
1,03 |
1,50 |
Маннит |
0,52 |
95,5 |
1,32 |
1,18 |
|
Фруктоза |
0,42 |
76,1 |
1,05 |
1,46 |
|
Глюкоза |
0,40 |
72,7 |
1,01 |
1,11 |
|
Candida utilis |
Глюкоза |
0,51 |
91,8 |
1,28 |
1,32 |
Penicillium chrysogenum |
Глюкоза |
0,43 |
77,4 |
1,08 |
1,35 |
Pseudomonas fluorescens Rhodopseudomonas spheroides |
Глюкоза |
0,38 |
68,4 |
0,95 |
0,85 |
Глюкоза |
0,45 |
81,0 |
1,12 |
1,46 |
|
Saccharomyces cerevisiae |
Глюкоза |
0,50 |
90,0 |
1,25 |
0,97 |
|
Aerobacter aerogenes |
Рибоза |
0,35 |
53,2 |
0,88 |
0,98 |
Сукцинат |
0,25 |
29,7 |
0,62 |
0,62 |
|
Глицерин |
0,45 |
41,8 |
1,16 |
0,97 |
|
Лактат |
0,18 |
16,6 |
0,46 |
0,37 |
|
Пируват |
0,20 |
17,9 |
0,49 |
0,48 |
|
Ацетат |
0,18 |
10,5 |
0,43 |
0,31 |
|
Candida utilis |
Ацетат |
0,36 |
21,0 |
0,90 |
0,70 |
Pseudomonas fluorescens |
Ацетат |
0,28 |
16,8 |
0,70 |
0,46 |
Candida utilis |
Этанол |
0,68 |
31,2 |
1,30 |
0,61 |
Pseudomonas fluorescens |
Этанол |
0,49 |
22,5 |
0,93 |
0,42 |
Klebsiella sp. |
Метанол |
0,38 |
12,2 |
1,01 |
0,56 |
Methylomonas sp. |
Метанол |
0,48 |
15,4 |
1,28 |
0,53 |
Pseudomonas sp. |
Метанол |
0,41 |
13,1 |
1,09 |
0,44 |
Methylococcus sp. |
Метан |
1,01 |
16,2 |
1,34 |
0,29 |
Pseudomonas sp. |
Метан |
0,80 |
12,8 |
1,06 |
0,20 |
Pseudomonas sp. |
Метан |
0,60 |
9,6 |
0,80 |
0,19 |
Pseudomonas methanica |
Метан |
0,56 |
9,0 |
0,75 |
0,17 |
а Воспроизведено с разрешения из работы: Nagai S., Mass and Energy Balances for Microbial Growth Kinetics, in Advances in Biochemical Engineering, vol. 11, Ghose T. K., Fiechter A., Blakebrough N. (eds.), Springer-Verlag, New York, p. 53, 1979.
В случае хемогетеротрофов один и тот же субстрат выполняет функции источника; энергии и углерода, поэтому суммарную утилизацию субстрата можно выразить следующим уравнением:
![]()
Разделив это уравнение на (∆X), получим
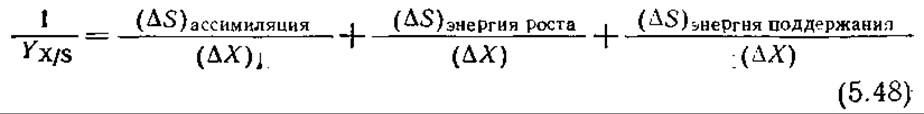
Истинный экономический коэффициент клеточного роста ∆Х/(∆S)ассимиляция относительно постоянен и представляет собой стехиометрически четко определенную величину. Этого, однако, нельзя сказать об общем коэффициенте Yх/s, что связано с зависимостью различных (∆S) в правой части уравнения (5.48) от условий роста. Так, в быстро растущей популяции клеток большая часть субстрата расходуется на ассимиляцию и рост, в то время как в популяции клеток, находящейся в стационарном состоянии или покое, субстрат часто расходуется исключительно на поддержание (в этом случае YX/S = 0).
В следующем разделе мы рассмотрим другие, более строгие стехиометрические выражения для клеточного роста с учетом уравнений материального баланса по углероду, водороду, азоту и кислороду. Кроме того, мы познакомимся с более детальным подходом к моделированию клеточного роста, при котором рост клеток все еще рассматривается как единый процесс, но вместе с этим учитывается ведущая роль NADH и АТР в метаболизме роста.