Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Стехиометрия и энергетика метаболических превращений
Стехиометрия клеточного роста и образования продуктов метаболизма
Стехиометрия фотосинтеза
Оценка выходов продуктов реакции фотосинтетического восстановления СО2 до углеводов важна как в процессах производства продуктов питания, так и для определения скоростей воспроизводства исходного сырья при получении биомассы. Известные сведения о последовательности реакций фотосинтеза и сопутствующих потоках электронов позволяют сделать вывод, что теоретически возможная эффективность фотосинтеза (выраженное в процентах отношение свободной энергии глюкозы к свободной энергии поглощенного излучения при 700 нм) составляет 35%. Конечно, эта величина представляет собой предельное значение, только теоретически достижимое в идеальных условиях. Здесь не учтены такие факторы, как утилизация в фотосинтезе только части солнечной радиации (соответствующий понижающий коэффициент равен приблизительно 0,43), отражение света и затенение листьев (понижающий коэффициент 0,80 или меньше), темновое дыхание (соответствующий коэффициент может изменяться в широких пределах; более или менее типичная величина составляет 0,67). Умножив все эти коэффициенты на теоретическую эффективность фотосинтеза, можно определить реальную (высшую) эффективность, составляющую около 8%.
Таблица 5.13. Максимальные значения выходов продуктов метаболизма по массе Ymaxp/s (выраженные в граммах продукта метаболизма на 1 г субстрата), вычисленные по уравнению (5.72)а
Субстрат |
|||
Продукт метаболизма |
н-Алканы |
Этанол |
Глюкоза |
Лимонная кислота |
4,55 |
2,77 |
1,42 |
Полисахариды |
3,21 |
1,96 |
1,00 |
Уксусная кислота |
3,21 |
1,96 |
1,00 |
Лизин |
2,23 |
1,36 |
0,70 |
Стрептомицин |
— |
— |
0,66 |
Этанол |
1,64 |
— |
0,51 |
Триглицериды |
1,18 |
0,72 |
0,37 |
а Воспроизведено с разрешения из работы: Eroshin V. К., Minkevich I. G., On the Upper Limit of Mass Yields of an Organic Product from an Organic Substrate, Biotech. Bioeng., 24, 2263 (5982).
Соответствующее количество биомассы (в виде крахмала), продуцируемое на единице площади (м2) в единицу времени (например, за 1 сут), можно оценить следующим образом:

Интенсивность солнечной радиации зависит от времени года и географической зоны. Например, средняя интенсивность солнечной радиации в США составляет 3930 ккал/(м2∙сут), а на юго-западе США в летний период достигает 6775 ккал/(м2∙сут). Последний сомножитель уравнения (5.73) учитывает потерю одной молекулы воды каждой молекулой глюкозы, включаемой в полимерную цепь крахмала.
Если имеются экспериментальные данные, характеризующие продуктивность процесса образования биомассы и интенсивность солнечной радиации, то по уравнению (5.73) можно рассчитать действительную эффективность фотосинтеза в растениях и водорослях (табл. 5.14). Найденные таким путем величины лежат в пределах от 0,8 до 3,2%, т. е. намного ниже теоретической оценки. Обратите внимание также на то, что эффективность в группе фотосинтезирующих так называемых С4-растений значительно выше, чем в группе С3-растений. Ниже мы вкратце рассмотрим биохимическую основу этого различия.
Таблица 5.14. Экспериментально найденные среднегодовые и максимальные значения эффективности и продуктивности фотосинтеза в некоторых растениях3
|
Растение |
Среднегодовые |
Найденные максимальные |
||
Выход, г/(м2∙сут) |
Эффективность, % |
Выход, г/(м2∙сут) |
Эффективность, % |
|
С3-Растения |
||||
Люцерна |
8 |
0,7 |
23 |
1,4 |
Сахарная свекла |
9 |
0,8 |
31 |
1,9 |
Хлорелла |
28 |
1,7 |
||
С |
||||
Сахарный тростник |
31 |
2,8 |
38 |
2,4 |
Сорго |
10 |
0,9 |
51 |
3,2 |
Кукуруза |
52 |
3,2 |
||
а Воспроизведено с разрешения из работы: Bassham J. A., Increasing Crop Production Through More-Controlled Photosynthesis: Science, 197, 630 (1977).
Явление фотодыхания (светового дыхания), т. е. поглощение кислорода и одновременное выделение СО2 на свету, представляет собой важный процесс, влияющий на общую эффективность роста фотосинтезирующих организмов. В этом процессе происходящие в растении реакции фиксации углерода приводят к промежуточному соединению — рибулозо-1,5-дифосфату (см. разд. 5.6.1), который далее распадается следующим образом:
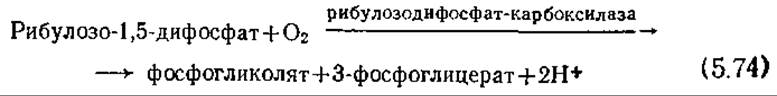
Затем гидролизуется фосфогликолят, образуя гликолят, который в свою очередь, в органоидах растительной клетки превращается в глицин и другие вещества.
В этом окислительном процессе не образуется АТР и не высвобождаются электроны; следовательно, здесь расходуется энергия фотосинтеза. В то же время реакция окисления (5.74) ингибируется высокими концентрациями СО2. В клетках С4-растений, в которых цикл Калвина (рис. 5.15) любопытным образом видоизменен, поддерживается относительно высокая концентрация СО2. В непосредственно контактирующих с воздухом клетках мезофилла С4-растений осуществляется ряд химических превращений, которые суммарно можно выразить следующим уравнением:
Клетки мезофилла:
Пируват + СО2 + АТР + 2Н2О → малат + АТР + 2Рі (5.75)
Образующийся таким образом малат диффундирует в соседние клетки обкладки проводящего пучка, в которых реализуются реакции цикла Калвина. Здесь включенный в малат СО2 выделяется в соответствии с реакцией
Малат → пируват + СО2 (5.76)
В конечном счете в этом метаболическом цикле для поддержания обеспечивающей минимум фотодыхания высокой концентрации СО2 вблизи от рибулозодифосфат-карбоксилазы расщепляются две высокоэпергетические фосфатные связи. Отсюда следует, что фотосинтез 1 моль глюкозы дополнительно требует 12 моль АТР. Таким образом в С4-растениях энергия, очевидно, расходуется на два конкурирующих процесса: цикл реакций транспорта СО2 и усиленное фотодыхание (в отсутствие цикла транспорта СО2). Интересно отметить, что энергия активации реакции фотодыхания (5.74) превышает энергию активации реакции карбоксилирования (5.37), которая также катализируется рибулозодифосфат-карбоксилазой. Возможно, именно этот факт может объяснить, почему С4-растения шире распространены в тропиках, а С3-растения преобладают в умеренной зоне.