Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Молекулярная генетика и системы регуляции
Модификация структуры клеточной ДНК
Трансформация и конъюгация в бактериях
Под трансформацией понимают процесс переноса генетической информации из одной клетки в другую в виде свободной ДНК. В этом процессе фрагмент двухцепочечной ДНК проникает в клетку-реципиент, находящуюся в специфическом состоянии, когда она способна включать чужеродную ДНК (такие клетки называют компетентными). Если перенесенный фрагмент аналогичен фрагменту ДНК реципиента, то хромосома последнего быстро трансформируется путем кроссинговера.
Механизм трансформации позволяет включать в живые бактерии не только фрагменты хромосом, но и плазмиды. В разд. 2.3 мы узнали, что плазмидой называется молекула ДНК, существующая и реплицирующаяся независимо от бактериальной хромосомы. Плазмиды обычно представляют собой относительно небольшие кольцевые молекулы с молекулярной массой порядка 106—108 (рис. 6.15). Трансформация с помощью плазмид является одним из основных приемов в методе направленного изменения структуры ДНК, рассматриваемом в разд. 6.3.
Плазмиды обычно не выполняют каких-либо существенных функций в нормальных условиях жизнедеятельности клетки, но в то же время могут обеспечивать клетку рядом полезных (в специфических условиях) качеств. Установлено, например, что плазмиды, называемые R-факторами (факторами устойчивости), обусловливают устойчивость бактерий к антибиотикам.
Плазмидам близки так называемые эписомы — молекулы ДНК, которые могут или быть составной частью клеточной хромосомы, или существовать независимо от нее. Один из наиболее известных примеров эписом — фактор F (фактор фертильности), характеризующий клетки Е. coli при их конъюгации. В процессе конъюгации Е. coli содержащие F--фактор клетки (F+-клетки) передают его клеткам, не имеющим фактора F (F--клеткам); иногда и часть хромосомы F+-клетки также передается F--клетке. Аналогичным образом могут передаваться и R-факторы. Впрочем, некоторые F+-мутанты, обозначаемые Hfr, ведут себя по-иному; они не инфицируют F--клетку F--фактором, а передают ей большую часть своей хромосомы, которая в результате кроссинговера с хромосомой F--клетки образует генетически модифицированные штаммы.
Многочисленными экспериментами показано, что хромосома F+-клетки проникает в F--клетку в линейном порядке в виде нити; если процесс прерывать через различные прохмежутки времени, то в F--клетках будут находиться участки хромосомы различной длины. Последующее сравнение характеристик популяций клеток-реципиентов, содержащих различные количества хромосомного материала из F+-клеток, позволяет выяснить расположение генетического материала в хромосоме F+-клегки. Именно таким путем были получены детальные генетические карты типа изображенной на рис. 6.16. На этой карте обозначены относительные положения характерных для различных мутантов генов. Например, символ thr указывает ген, характерный для мутанта, который растет только в присутствии треонина. Расшифровка многих подобных обозначений приведена в монографии Уотсона (ссылка [7] в гл. 2).
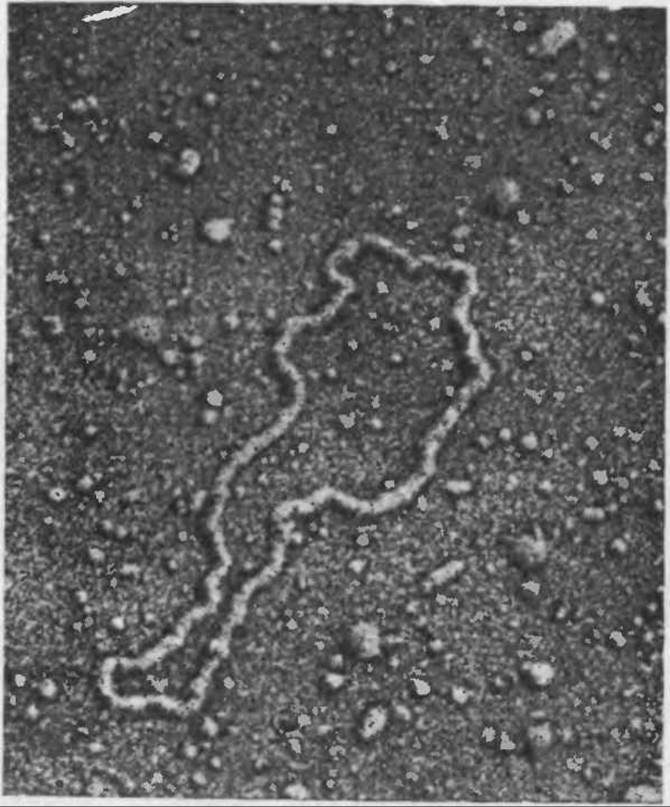
РИС. 6.15. Электронная микрофотография плазмиды pSC101 (Х230 000). Эта кольцевая ДНК, существующая и реплицирующаяся независимо от бактериальной хромосомы, была важным инструментом в первых экспериментах с рекомбинантными ДНК. (Микрофотография предоставлена д-ром С. Н. Коуэном.)