Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Молекулярная генетика и системы регуляции
Технология рекомбинантных ДНК
Ферменты для расщепления и сшивки ДНК
Успехи генетической инженерии во многом связаны с выделением и последующим промышленным производством ряда ферментов, с помощью которых можно разрезать, видоизменять и соединять молекулы ДНК в лабораторных условиях. Особое место среди этих ферментов занимают рестрикционные эндонуклеазы, способные узнавать и расщеплять специфические нуклеотидные последовательности в молекулах ДНК. Например, рестриктаза ЕсоRI специфична по отношению к последовательности шести нуклеотидных остатков в двойной спирали следующего типа:

Эта рестриктаза расщепляет каждую цепь между остатками А и G по связям, отмеченным стрелками. В результате образуются два фрагмента:
![]()
Обратите внимание на то, что каждый фрагмент содержит короткую одноцепочечную последовательность, называемую «липким концом», поскольку эти две последовательности комплементарны и, следовательно, имеют тенденцию к связыванию друг с другом за cчет водородных связей между парными основаниями. Реализация этой тенденции зависит от экспериментатора, который посредством изменения pH или температуры может добиться образования водородных связей или их разрушения.
К настоящему времени идентифицированы более 100 различных ферментов, обладающих рестриктазной активностью. В табл. 6.7 перечислены наиболее часто используемые рестриктазы, соответствующие сайты узнавания и гидролизуемые ими фосфодиэфирные связи. Нетрудно заметить, что не все рестриктазы образуют липкие концы. Рестриктаза HpaІ, например, расщепляет специфическую последовательность шести нуклеотидных остатков таким образом, что образуются только двухцепочечные фрагменты с «тупыми концами».
Таблица 6.7. Некоторые рестрикционные эндонуклеазы, соответствующие сайты узнавания и расщепляемые связи (указаны стрелками)
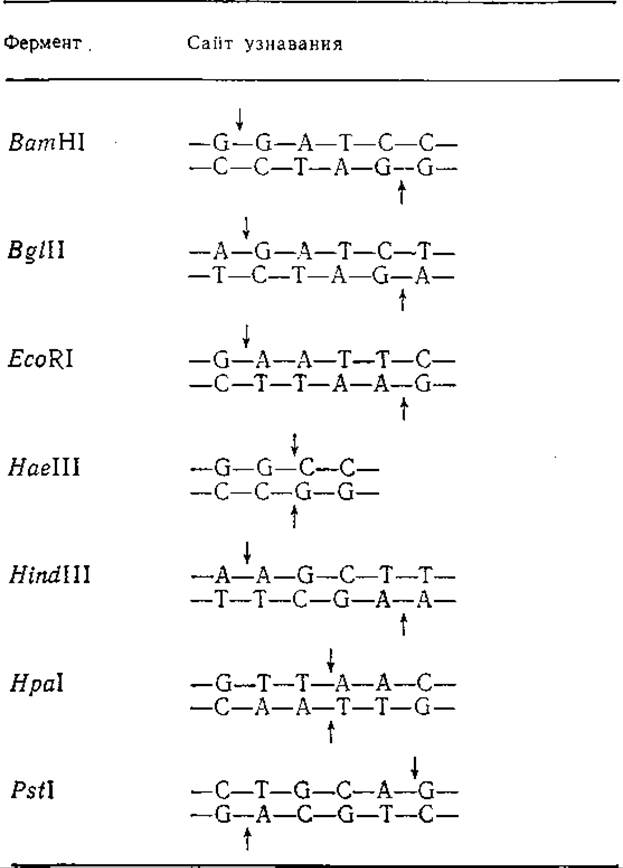
Номенклатура рестрикционных ферментов базируется на названии организма, из которого данная рестриктаза была впервые выделена. Первая прописная буква в обозначении рестриктазы соответствует первой букве названия рода организма, а последующие две или три строчные буквы берутся из первых двух или трех букв названия вида этого организма. Наконец, римская цифра обозначает порядковый номер фермента, отражающий хронологию его выделения из данного организма. Так, рестриктаза BglII представляет собой вторую рестриктазу, обнаруженную в бактерии Bacillus globigii. Для уточнения природы штамма, продуцирующего данный фермент, в название рестриктазы иногда включают дополнительные буквы; так, рестриктаза EcoRI была выделена из Е. coli RY13.
Ценность рестрикционных ферментов в технологии рекомбинантных ДНК обусловливается их высокой специфичностью, т. е. способностью расщеплять только строго определенные нуклеотидные последовательности. Поскольку вероятность наличия в молекуле ДНК специфических сайтов узнавания, содержащих от 4 до 6 нуклеотидных остатков, не очень велика, то образующиеся в результате гидролиза рестриктазами фрагменты ДНК обычно имеют довольно большую молекулярную массу (как правило, несколько сотен нуклеотидных остатков). Это достаточно много для того, чтобы образовавшиеся фрагменты несли ценную генетическую информацию, и одновременно довольна мало для того, чтобы их можно было изучать и модифицировать in virto.
Объем настоящего учебного пособия позволяет лишь вкратце упомянуть другие важные ферменты, применяющиеся в технологии рекомбинантных ДНК. В первую очередь следует отметить ДНК-лигазу. Предположим, что мы создали такие условия, когда липкие концы, образовавшиеся в результате расщепления различных молекул ДНК под действием EcoRI, «отжигаются», т. е. сближаются и связываются за счет образования пар оснований. Образовавшиеся ДНК не будут, однако, ковалентно замкнутыми, поскольку между остатками А и G в обеих цепях отсутствуют фосфодиэфирные связи. Последние могут быть вновь созданы с помощью ДНК-лигазы, катализирующей конденсацию 3'-гидроксильной группы с 5'-фосфатной группировкой.
Таким путем, например, отожженные фрагменты
![]()
ковалентно сшиваются ДНК-лигазой:
![]()
В заключение отметим другие ферменты, катализирующие синтез олигонуклеотидов (рис. 6.22). На матричной цепи ДНК фермент ДНК-полимераза катализирует построение комплементарной цепи в 5'—3'-направлении. Для осуществления этого процесса необходимо присутствие всех четырех дезоксирибонуклеотидов, а также праймера, обладающего свободной 3'-гидроксильиой группой. Обратная транскриптаза (ревертаза; РНК-зависимая ДНК-полимераза) выполняет задачу, потребность в решении которой возникает сравнительно редко; этот фермент как бы обращает обычный порядок передачи генетической информации, синтезируя цепь ДНК, комплементарную матрице мРНК. Соединение различных фрагментов ДНК часто облегчается после их удлинения на комплементарные гомополимерные цепи (хвосты); в присутствии только одного нуклеотидтрифосфата фермент концевая трансфераза способен многократно присоединять этот нуклеотид к концевой 3'-ОН-группе молекулы ДНК.
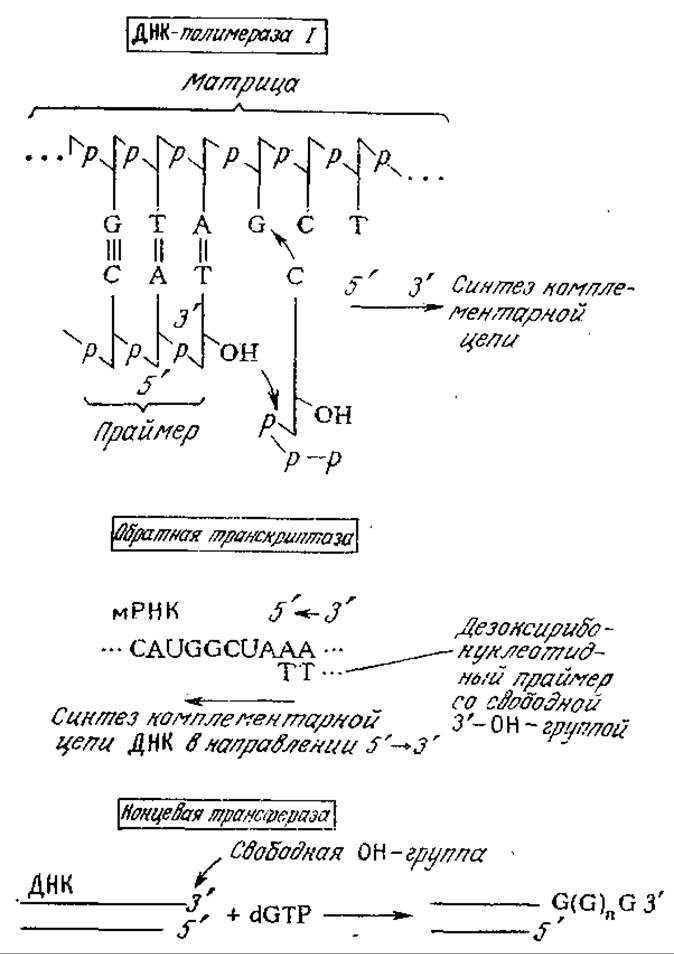
РИС. 6.22. Характерные реакции некоторых синтезирующих ферментов, используемых при получении рекомбинантных ДНК.
Некоторые другие ферменты, играющие большую роль в методологии генетической инженерии, мы упомянем ниже, а следующий раздел будет посвящен требованиям, предъявляемым к векторам рекомбинантных ДНК.