Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Кинетика процессов утилизации субстрата, образования продуктов метаболизма и биомассы в культурах клеток
Кинетика сбалансированного роста
Влияние эндогенного метаболизма и метаболизма поддержания на кинетику клеточного роста
Приведенные на рис. 7.8 данные для культуры A. aerogenes показывают, что при малых скоростях разведения концентрация клеток заметно снижается. Аналогичное поведение наблюдалось и в случае кормовых дрожжей Torula utilis. Эту особенность, противоречащую модели Моно для хемостата, можно объяснить, если в модели учесть возможность эндогенного метаболизма. Под эндогенным метаболизмом подразумеваются происходящие в клетке реакции, в которых расходуются клеточные вещества. Чтобы учесть этот эффект, дополним уравнение Моно слагаемым —kex:
![]()
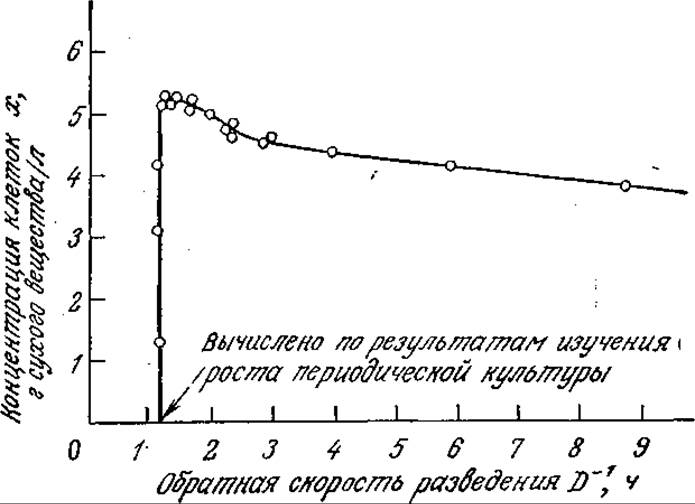
РИС. 7.8. Снижение концентрации клеточной массы (культура A. aerogenes) в глицериновой среде в проточном реакторе при уменьшении скорости разведения противоречит модели Моно. (Воспроизведено из работы: Herbert D., Continuous Culture of Microorganisms: Some Theoretical Aspects, in Continuous Culture of Microorganisms: A Symposium, p. 48, Publishing House of the Czechoslovakia Academy of Sciences, Prague, 1958.)
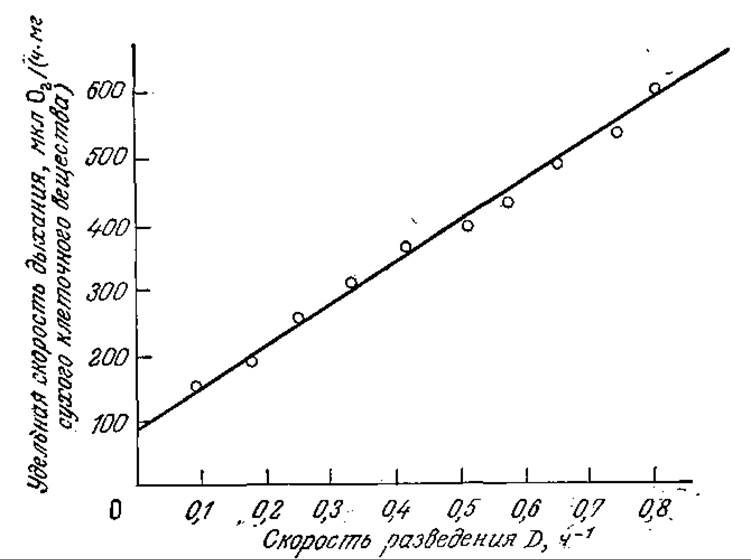
РИС. 7.9. Согласующаяся с уравнением (7.24) линейная зависимость удельной скорости дыхания бактерии А. aerogen.es от скорости разведения в проточном реакторе. (Воспроизведено из работы: Herbert D., Continuous Culture о! Microorganisms: Some Theoretical Aspects, in Continuous Culture of Microorganisms: A Symposium, p. 49, Publishing House of the Czechoslovakia Academy of Sciences, Prague, 1958.)
Обратите внимание на то, что слагаемое — kex в уравнении (7.22) можно интерпретировать и как скорость гибели клеток.
Модифицированное уравнение Моно согласуется и с другими экспериментальными данными. Например, если скорость дыхания аэробной культуры пропорциональна скорости утилизации субстрата, т. е.
![]()
то из уравнений (7.8), (7.22) и (7.23) следует, что удельная скорость дыхания равна
![]()
Приведенные на рис. 7.9 экспериментальные данные хорошо согласуются с уравнением (7.24).
Наблюдаемая зависимость экономического коэффициента Y от D также подтверждает справедливость уравнения (7.22) для скорости клеточного роста. Если скорость поглощения субстрата равна
![]()
где Y'x/s — «истинный» коэффициент, то из уравнений (7.22) и (7.25) и определения Yх/s (напомним, что Yх/s — это стехиометрический коэффициент, равный частному от деления общей массы образовавшихся клеток на общую массу поглощенного субстрата, поэтому при условии стерильности питательных веществ -rs = Dx/Yх/s) следует, что
![]()
Экспериментально показано, что для ряда микроорганизмов зависимость Yx/s от D выражается именно таким уравнением.
Другая возможность, рассмотренная вкратце при обсуждении стехиометрии клеточного роста в разд. 5.10.1, заключается в параллельной утилизации субстрата как для клеточного роста, так и для других энергетических потребностей клетки (метаболизм поддержания). В этом случае скорость утилизации субстрата определяется выражением
![]()
где m — удельная скорость потребления субстрата, расходуемого в метаболизме поддержания. Если принять, что rх равно μх, то
![]()
Таким образом мы получили точно такую же функциональную зависимость Yx/s от D, как и в случае модели эндогенного метаболизма [уравнение (7.26)].
Как показано на рис. 7.10, при высоких скоростях разведения поведение культуры клеток в проточном реакторе может существенно отклоняться от поведения, предсказываемого моделью Моно для хемостата. Здесь не только слишком велика ошибка в определении концентрации клеток вблизи точки вымывания, но и вся популяция клеток может существовать при скоростях разведения, значительно превышающих предсказываемую теорией критическую величину. Более того, по мере приближения D к критическому максимальному значению экономический коэффициент снижается. Одна из возможных причин этого несоответствия может заключаться в относительно высокой концентрации субстрата, характерной для высоких скоростей разведения. В таких условиях субстрат часто не ограничивает рост клеток, и составляющие популяцию клетки могут изменить характер метаболизма, выбрав какой-либо другой лимитирующий фактор среды. Еще одной причиной может быть недостаточно эффективное перемешивание; этот вопрос мы изучим в гл. 9.
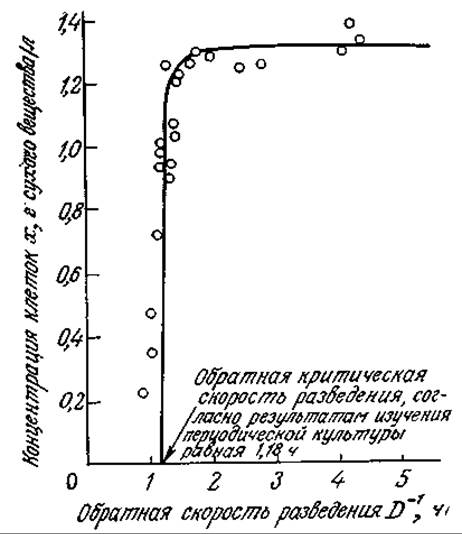
РИС. 7.10. Экспериментальные данные по росту Aerobacter cloacae в проточном реакторе, свидетельствующие об отклонении концентрации клеточной массы от нулевого значения при скоростях разведения, превышающих вычисленную критическую величину. [Воспроизведено из статьи: Herbert D. et al., The Continuous Culture of Bacteria: A Theoretical and Experimental Study, J. Gen. Microbiol., 14, 601 (1956).]