Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Процессы переноса в биотехнологических системах
Массообмен между газовой и жидкой фазами в клеточных системах
Скорость утилизации кислорода в процессах клеточного метаболизма
Проектируя аэробные биологические реакторы, мы с помощью определенных математических выражений, с большим или меньшим приближением описывающих интересующую нас ситуацию, часто пытаемся выяснить, является ли самой медленной стадией процесса перенос кислорода (или другого лимитирующего клеточный рост вещества) или его утилизация клетками. Максимально возможная скорость массообмена определяется путем простой подстановки в указанные выше уравнения сl = 0; при этом условии весь поступающий в раствор кислород быстро усваивается клетками. В предыдущей главе мы установили, что максимальная скорость утилизации кислорода составляет xμmах/YO2, где х — плотность культуры клеток, а YO2 — отношение числа грамм-атомов углерода в образующихся клеточных углеродсодержащих соединениях к числу молей усвоенного кислорода.
Если kla'cl* намного больше xμmах/YO2, то, очевидно, основным препятствием дальнейшему повышению скорости утилизации кислорода является метаболизм микроорганизмов и скорость процесса ограничивается биохимическими факторами. В другом крайнем случае, когда kla'cl* ≪ xμmax/YO2, кажущееся значение cl приближается к нулю и скорость процесса в реакторе должна определяться явлениями переноса.
В действительности ситуация несколько сложнее. В стационарном состоянии скорости утилизации и абсорбции кислорода должны быть равны
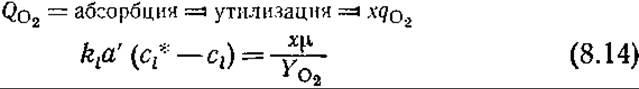
Если известна зависимость μ от cl, то с помощью уравнения (8.14) можно определить cl и, следовательно, скорость утилизации кислорода.
В общем случае существует некоторая критическая концентрация кислорода в жидкой фазе, при достижении которой все клетки насыщаются О2 и количество последнего превышает минимум, необходимый для немедленного захвата всех электронных пар, участвующих в реакциях дыхательной цепи; в этой ситуации, следовательно, скорость процесса будет лимитировать какой-то иной биохимический механизм (гл. 5). Например, если зависимость удельной скорости роста клеток μ от концентрации кислорода подчиняется уравнению Моно, то
![]()
Общее решение уравнений такого типа было дано в разд. 4.4.1, но здесь для наглядности примем, что величина cl значительно меньше cl*. Для биологических реакторов такая ситуация довольно обычна. Если cl ≪ cl*, то cl можно выразить следующим образом:
![]()
Если найденная таким путем величина cl больше критического значения cсr (около 3KO2), то скорость утилизации кислорода микроорганизмами лимитируется каким-либо иным фактором, например низкой концентрацией другого субстрата, даже если средняя концентрация растворенного кислорода в жидкой фазе значительно ниже уровня насыщения. Величины критических концентраций кислорода для различных организмов лежат в диапазоне от 0,003 до 0,05 ммоль/л (табл. 8.2) или от 0,1 до10% максимальной растворимости O2 (табл. 8.1), или от 0,5 до 50% конц. насыщения кислородом воздуха. Очевидно, что массообмен кислорода особенно важен в случае организмов, для которых характерны большие значения критической концентрации кислорода, например плесеней Penicillium.
Таблица 8.2. Типичные значения сО2,сr в присутствии субстратаa
Организм |
Температура, °С |
cO2, cr, ммоль/л |
Azotobacter vinelandii |
30 |
0,018-0,049 |
Е. coli |
37,8 |
0,0082 |
15 |
0,0031 |
|
Serratia marcescens |
31 |
~0,015 |
Pseudomonas denitrificans |
30 |
~0,009 |
|
Дрожжи |
34,8 |
0,0046 |
20 |
0,0037 |
|
|
Penicillium chrysogenum |
24 |
~0,022 |
30 |
~0,009 |
|
Aspergillus oryzae |
30 |
~0,020 |
а Из работы: Finn R. К., р. 81 in Biochemical and Biological Engineering Science, Blakebrough N. (ed.), vol. 1, Academic Press, Inc., New York, 1967.
Различные факторы могут влиять на общую потребность микроорганизма в кислороде хμ/YO2, которая в свою очередь определяет необходимые в проектировочных работах минимальные значения kla' [уравнение (8.14)]. К числу наиболее важных факторов такого типа относятся вид клеток, фаза роста культуры, источники углерода, pH среды, тип изучаемого микробиологического процесса; последний фактор включает параметры утилизации субстрата, образования биомассы или выхода продукта жизнедеятельности микроорганизмов (гл. 7).
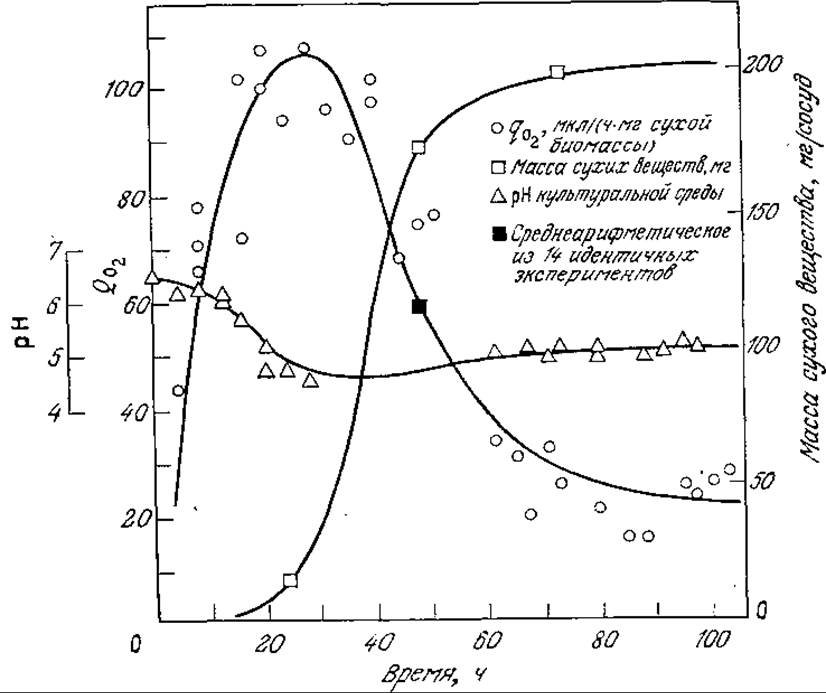
РИС. 8.5. Скорость утилизации кислорода периодической культурой Myrothecium verrucaria. [Воспроизведено из статьи: Darby R. Т., Goddard D. R., Аm. J. Bot., 37, 379 (1950).]
В приведенных на рис. 8.5 результатах определения скорости утилизации кислорода в периодическом процессе удельная потребность в О2 достигает максимума в начале экспоненциальной фазы, а х возрастает и на более поздних этапах роста. Произведение хμ, а следовательно, и общая потребность в кислороде максимальны в конце экспоненциальной и в самом начале стационарной фазы; таким образом, скорость утилизации кислорода достигает максимума позднее, чем удельная скорость роста.
На скорость утилизации кислорода большое влияние оказывает природа углерод содержащего питательного вещества. Глюкоза, например, в общем случае метаболизируется быстрее, чем другие углеводы. Найдено, что максимальная скорость потребления кислорода Penicillium составляет 4,9, 6,7 и 13,4 моль/(л∙ч) в случае роста плесени на лактозе, сахарозе и глюкозе соответственно [2].
Суммарная потребность клетки в кислороде складывается из потребностей на поддержание жизнедеятельности клеток, окислительные реакции в процессе дыхания (используемые для роста клеток и для целей биосинтеза) и окисление субстратов в соответствующие конечные продукты метаболизма. При изучении стехиометрии метаболизма (гл. 5) мы видели, что скорость утилизации кислорода, расходуемого на клеточный рост, обычно непосредственно связана с количеством потребляемого субстрата, являющегося источником углерода. Кроме того, более восстановленные субстраты, например метан и высшие углеводороды, требуют усиленной утилизации кислорода клетками по сравнению с субстратами типа глюкозы, степень восстановленности атомов С в которых приблизительно та же, что и в компонентах клетки. Так, коэффициенты YO2/с [т. е- отношение числа молей утилизированного кислорода к числу молей метаболизированного субстрата (в расчете на одноуглеродный субстрат)] равны 1,34, 1,0 и 0,4 для типичных микроорганизмов, растущих на метане, парафиновых углеводородах и углеводах соответственно.
Кислород может также выполнять роль реагента, участвующего в биотрансформациях. В производстве 5-кетоглюконовой кислоты из глюкозы, например, на первой стадии роста периодической культуры Acetobacter происходит окисление части содержащейся в среде глюкозы до глюконовой кислоты; при этом О2 используется как для клеточного роста, так и для синтеза продукта метаболизма. После истощения глюкозы рост клеток прекращается, а глюконовая кислота превращается в 5-кетоглюконовую кислоту по следующему уравнению реакции:
С6Н12О7 + 1/2O2 → С6Н10О7 + Н2О (8.17)
В заключительной стадии процесса осуществляется только указанная реакция, и потребление кислорода связано с образованием продукта метаболизма в соответствии со стехиометрией уравнения (8.17).