Основы биохимии - Филиппович Ю. Б. 1999
Общее понятие об обмене веществ и энергии в организме
Характеристика структуры, свойств и функциональной активности ферментов, коферментов, витаминов и ряда других биологически активных соединений, приведенная в предыдущих главах, неизбежно сопровождалась рассмотрением тех биохимических процессов, в которых они участвуют. Этим невольно был сделан первый шаг к изучению некоторых аспектов обмена веществ. В последующих главах это важнейшее свойство живого будет освещено во всех его многообразных проявлениях. Здесь же целесообразно сосредоточить внимание на общих закономерностях.
Общие представления об обмене веществ. Обмен веществ и энергии составляет сущность жизнедеятельности любого организма. Для самого явления жизни характерен постоянный обмен веществ с окружающей внешней природой. Живое остается живым до тех пор, пока оно способно строить самое себя из веществ окружающей среды; живая материя поддерживает собственное существование путем постоянного и непрерывно протекающего с той или иной скоростью поглощения химических соединений из внешнего по отношению к ней мира, преобразования их в конституционные элементы своего тела или более простые соединения и, наконец, выведения во внешнее пространство продуктов распада как собственного тела, так и преобразованных в процессе жизнедеятельности веществ. Этот непрерывный, самосовершающийся и саморегулируемый круговорот веществ, протекающий в процессе существования живой материи и сопровождающийся ее постоянным самообновлением, называется обменом веществ.
Иначе говоря, обмен веществ есть закономерный, самосовершающийся процесс превращения материи в живых телах.
Преобразование химических соединений, взаимодействие их друг с другом, разрушение одних и построение других происходит и в неживой материи. Однако при этом неживая система любого типа не самообновляется, а лишь видоизменяется. Жизнь, живая материя возникли тогда, когда среди неживых систем с характерными для них процессами неупорядоченного изменения веществ в природе появились системы, способные поддерживать собственное существование путем упорядоченного преобразования и использования факторов окружающей среды. Поэтому только та форма материи, те системы, которые обладают обменом веществ, получили название живой материи.
Явление жизни необычайно сложно и многогранно. «Как часть планетного земного живого вещества мы инстинктивно и бессознательно ярко чувствуем загадку жизни — своего существования и существования жизни», — писал академик В. И. Вернадский1.
1 Вернадский В. И. Химическое строение биосферы Земли и ее окружения. М., 1987.
В связи со значительными успехами квантовой биохимии, молекулярной биологии, кибернетики, термодинамики и генетики в последнее время предпринято много попыток углубить и конкретизировать понимание сущности жизни. С точки зрения квантовой биохимии жизнь определяют как форму динамической реализации квантовых характеристик атомов. В молекулярной биологии приходят к обобщению, согласно которому жизнь представляется как макромолекулярная иерархическая организация, характеризующаяся репликацией, метаболической цикличностью и отточенной регуляцией энергетических процессов. При этом большая роль в явлениях жизни отводится нуклеиновым кислотам. В кибернетике особое внимание уделяют процессам выработки и передачи информации, характеризуя жизнь как высокоустойчивое состояние вещества, использующее для выработки сохраняющих реакций информацию, кодируемую состоянием отдельных молекул. С точки зрения термодинамики наиболее существенным свойством живых организмов является устойчивая термодинамическая неравновесность и способность поддерживать ее на определенном уровне. Если неживая система стремится прийти в равновесие с окружающей средой, то живая, напротив, старается уйти от этого равновесия, целенаправленно поддерживая свое неравновесное по отношению к окружающей среде состояние. Исходя из этого, жизнь определяют как способ существования динамически самосохраняющейся формы материи — живой природы, сущность которого сводится к грандиозному процессу циклической трансформации органического вещества, основанному на взаимодействии синтеза и распада живой протоплазмы, т. е. биотическом круговороте. В этом же аспекте жизнь определяют как потенциально способную к самовоспроизведению открытую систему сопряженных органических реакций, катализируемых последовательно и почти изотермично сложными и специфическими органическими катализаторами, которые сами вырабатываются этой системой.
Для современного генетика жизнь на Земле вырисовывается как интегральное существование ДНК, РНК и белков в форме индивидуализированных личных и видовых целостных структурно-биохимических саморегулирующихся открытых систем, со свойствами воспроизведения исторически развивающихся форм генетической информации. При оценке сущности жизни с позиций эволюционного учения на первый план выступают происхождение, организация и взаимодействие определенных надмолекулярных и клеточных структур, закономерности возникновения их совокупности в виде самостоятельной биологической единицы. По мнению физико-химиков, жизнь есть динамическое равновесие многофазных систем. Все перечисленные подходы к более глубокому познанию сущности жизни безусловно заслуживают внимания и каждый из них раскрывает новые стороны этого сложнейшего явления, но каждый из них в отдельности односторонен.
По мере изучения жизни наши представления о роли отдельных, конкретных химических соединений, равно как и о значении тех или иных процессов в ее осуществлении, будут неизбежно изменяться. Кроме того, выход человечества в Космос делает вероятным знакомство с неземными формами жизни, вещественная и функциональная характеристики которых могут оказаться в каждом случае уникальными. Поэтому определение жизни как особой, качественно отличной биологической формы движения материи, возникающей на определенном этапе ее развития, является, видимо, наиболее фундаментальным и удачным.
Биологическая форма движения материи характеризуется существованием саморегулирующихся и самовоспроизводящихся систем, обменивающихся с окружающей средой веществом и энергией. Поскольку закономерное преобразование веществ лежит в основе как процесса саморегуляции, так и процесса самовоспроизведения и условием их осуществления является непрерывное поступление одних веществ в систему и выведение других из нее, обмен веществ является важнейшим элементом жизни.
Та часть общего процесса обмена веществ, которая выражается в поглощении, накоплении, усвоении организмом веществ окружающей среды и создании, синтезе за их счет структурных единиц своего тела, называется анаболизмом или ассимиляцией. Та часть общего процесса обмена веществ, которая состоит в разрушении веществ, составляющих организм, в распаде элементов живого тела и выведении продуктов этого распада из организма, называется катаболизмом или диссимиляцией. Следовательно, обмен веществ есть диалектическое единство противоположных процессов питания и выделения, усвоения и разрушения, синтеза и распада, иными словами — диалектическое единство процессов ассимиляции и диссимиляции.
Обмен веществ представляет собой сочетание многих разнообразных и противоположных процессов. Одни из них представляют процессы физиологические (питание, выделение и т. п.), другие — физические (сорбция, перенос и т. п.), третьи — химические (распад, синтез и т. п.). Та часть процессов обмена веществ в организме, которая заключается в осуществлении химических реакций, ведущих к преобразованию индивидуальных химических соединений при их распаде и синтезе в процессе жизнедеятельности организма, называется промежуточным обменом веществ или метаболизмом. Именно промежуточный обмен веществ изучает динамическая биохимия.
Масштабы обмена веществ в живой природе колоссальны. Биомасса Земли, составляющая, по подсчетам ученых, от 1,8 ∙ 1012 до 2,4 ∙ 1012 т (в пересчете на сухое вещество), непрерывно обновляет свой состав, поглощая и выделяя огромные количества химических веществ. По подсчетам А. А. Ничипоровича, растения Земли за год усваивают из атмосферы около 650 млрд, т СО2 и выделяют в атмосферу около 350 млрд, т О2. За этот же срок растения извлекают из почвы около 5 млрд, т N, около 1 млрд, т Р и 10—15 млрд, т. других минеральных элементов, образуя около 380 млрд, т биомассы (в расчете на сухое вещество). Только свободно живущие в почве азотфиксирующие микроорганизмы и клубеньковые бактерии ежегодно связывают около 100 млн. т молекулярного N из воздуха. За 2000 лет весь О2 атмосферы Земли проходит через живое вещество. Выделение кислорода в результате фотосинтеза, протекающего в растениях, водорослях и фотосинтезирующих бактериях, составляет от 2 тыс. до 5 тыс. т в одну секунду. Ежедневно на Земле разрушается до СО2 и Н2О около 1 млрд, т органических соединений. По данным В. А. Ковды, полное обновление биомассы суши происходит в течение 200 лет. По мнению В. И. Вернадского, все вещество биосферы в течение краткого в геологических масштабах периода может пройти через живые организмы. Таким образом, «...жизнь — живое вещество — поистине является одной из самых могущественных геохимических сил нашей планеты, а вызываемая ею биогенная миграция атомов представляет форму организованности первостепенного значения в строении биосферы»1.
1 Вернадский В. И. Химическое строение биосферы Земли и ее окружения. М., 1987.
Если перейти от рассмотрения обмена веществ в масштабе живого вещества всей планеты к оценке его роли в мельчайшей единице живого — клетке, то здесь еще более ярко выступает значение обменных процессов в явлениях жизни. Клетка, имеющая согласно современным представлениям (см. рис. 3) сложнейшую внутреннюю организацию, является средоточением нескольких тысяч различных веществ. В простейшей бактериальной клетке (см. табл. 1) находится около 300 млн. молекул органических соединений примерно 5000 наименований и громадное число молекул воды и неорганических солей. Они не только закономерно расположены в клеточном объеме, но и находятся в постоянном физическом и химическом движении. Первое осуществляется благодаря наличию в клетке большого количества протоков, соединяющих различные ее части друг с другом. Второе состоит в непрерывном преобразовании химических веществ посредством многочисленных реакций распада и синтеза белков, нуклеиновых кислот, углеводов, липидов и т. п. Все эти реакции ускоряются сотнями ферментов и осуществляются согласованно во времени и пространстве, а самое главное — они совершаются с огромными скоростями и саморегулируются.
Обмен веществ осуществляется при условии и в результате постоянного взаимодействия живой и неживой материи, организма и среды. Естественно поэтому, что ход обмена веществ в организме, а часто и сам характер этого обмена находятся в тесной зависимости, в единстве с условиями внешней среды. Присущие организмам молекулярные механизмы видоизменения, преобразования, воспроизводства и разрушения специфических органических соединений (нуклеиновые кислоты, белки, липиды, углеводы и др.) действуют лишь в определенных, ограниченных интервалах температуры, давления, радиации. Они осуществляются лишь при условии постоянного притока веществ, пригодных для преобразования, и оттока веществ, которые уже не могут служить исходным материалом для построения тела данного организма.
Более того, поддержание на известном уровне специфической активности молекулярных механизмов, преобразующих вещества клетки, зависит от непрерывного воспроизведения составных частей самих этих механизмов в едином процессе обмена веществ. В этом смысле любой организм представляет неизбежно самонастраивающуюся, саморегулирующуюся систему, закономерно (и адекватно) изменяющуюся при изменении условий среды, с которыми организм взаимодействует. Таким образом, тип обмена веществ складывается в процессе жизнедеятельности организма как единство внутренних (консервативных) и внешних (изменчивых) факторов. В этом и состоит диалектическое единство организма и среды.
Закономерности обмена энергии. Обмен веществ невозможен без сопутствующего ему обмена энергии. Каждое органическое соединение, входящее в состав живой материи, обладает определенным запасом потенциальной энергии, за счет которой может быть совершена работа. Эту энергию принято называть свободной энергией. Уровни свободной энергии индивидуальных исходных веществ и продуктов реакции, как правило, различны, вследствие чего в процессе преобразования веществ происходит перераспределение свободной энергии между компонентами реакционной смеси, т. е. протекает обмен энергией между веществами.
Понятие о макроэргических соединениях. Главными материальными носителями свободной энергии в органических веществах являются химические связи между атомами, поэтому при преобразовании химических связей в молекуле уровень свободной энергии соединения изменяется. Если изменение уровня свободной энергии соединения при возникновении или распаде химической связи составляет около 12,5 кДж/моль преобразуемого вещества, то такая связь по своему энергетическому уровню считается нормальной. Именно такую размерность имеет изменение уровня свободной энергии при преобразовании большинства связей в органических соединениях. Однако при новообразовании и распаде некоторых связей уровень свободной энергии в молекулах ряда органических соединений изменяется в гораздо большей степени и составляет 25—50 кДж/моль и более. Такие соединения называются макроэргическими соединениями, а связи, при преобразовании которых наступают столь крупные изменения в энергетическом балансе вещества, — макроэргическими связями. Последние в отличие от обычных связей обозначают значком ~. Ниже приведены некоторые органические вещества первого (с нормальными связями) и второго (с макроэргическими связями) типов. В каждом случае в скобках указаны величины уменьшения свободной энергии (отмечено знаком минус перед цифрами в скобках) в исходных веществах по сравнению с таковыми в продуктах их гидролиза (при pH 7, t = 25° С, кДж/моль):
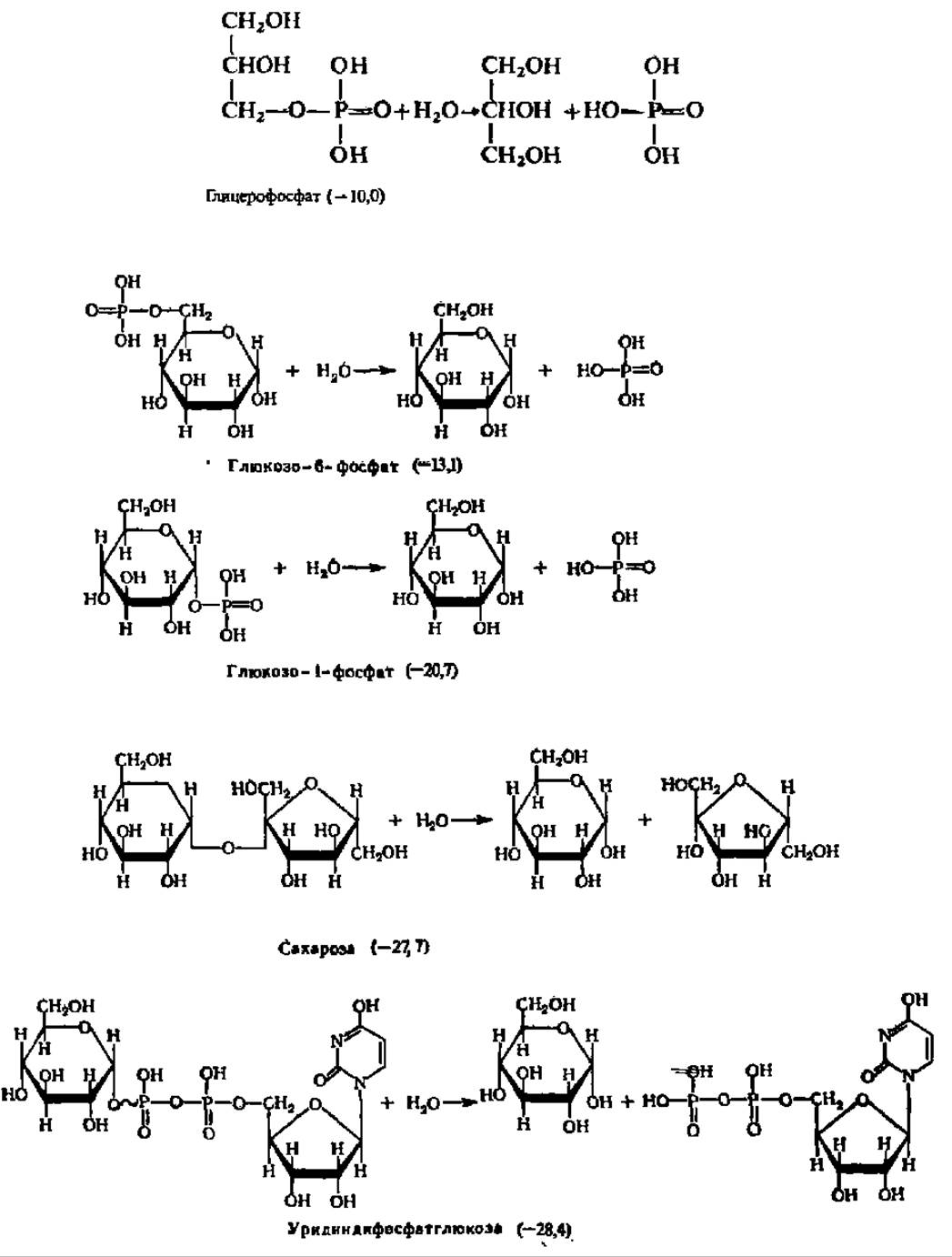
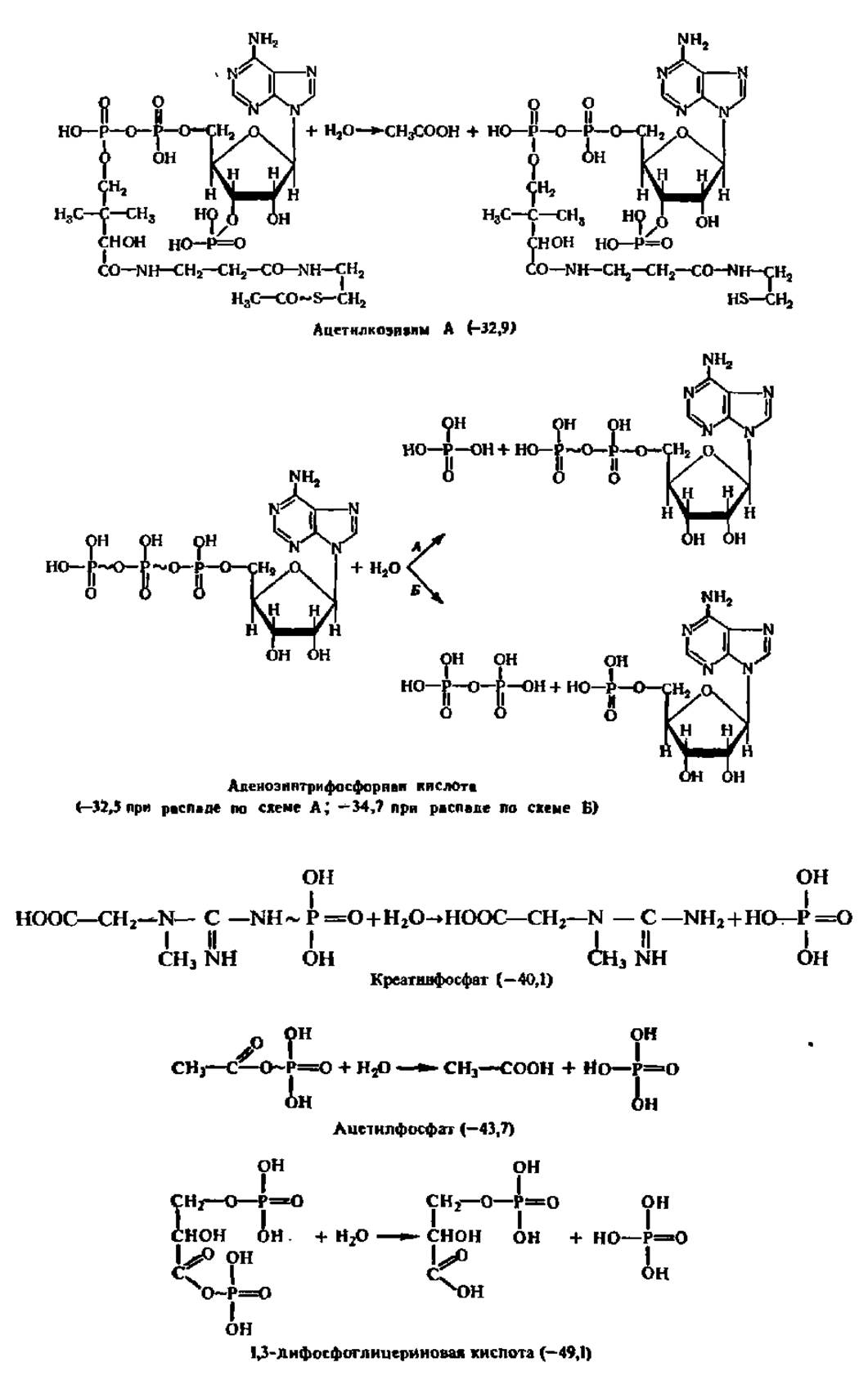
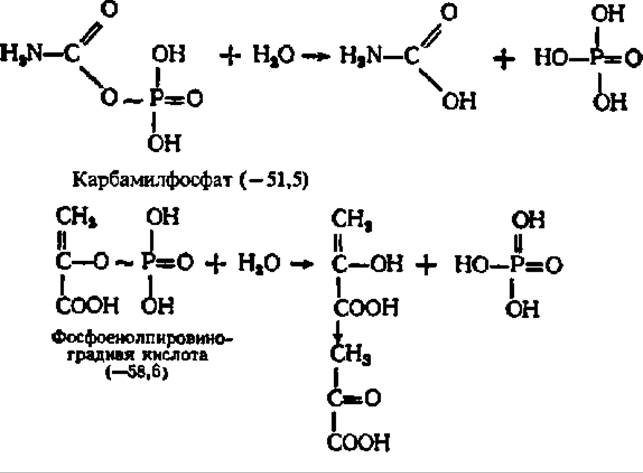
Нетрудно видеть, что макроэргические связи представлены преимущественно сложноэфирными, в том числе и тиоэфирными, ангидридными и фосфоамидными связями. Однако наиболее интересно, что почти все известные соединения с макроэргическими Связями содержат атомы Р и S, по месту которых в молекуле эти связи локализованы. Особая роль атомов S и Р в образовании макроэргических связей объясняется следующими причинами: 1) химические связи, свойственные элементам III периода, слабее, чем связи, характерные для элементов II периода; 2) S и Р образуют более четырех ковалентных связей; 3) только S и Р в III периоде сохраняют способность к замыканию кратных связей.
Именно та энергия, которая высвобождается при разрыве макроэргических связей, поглощается при синтезе органических соединений с более высоким уровнем свободной энергии, чем исходные. В то же время запасы макроэргических веществ в организме постоянно пополняются путем аккумулирования энергии, выделяющейся при понижении энергетического уровня распадающихся соединений.
Таким образом, макроэргические вещества выполняют функцию и доноров, и акцепторов энергии в обмене веществ; они служат как аккумуляторами, так и проводниками энергии в биохимических процессах. Кроме того, им свойственна роль трансформаторов энергии, так как они способны преобразовывать стационарную форму энергии химической связи в мобильную, т. е. в энергию возбужденного состояния молекулы.
Последний вид энергии и служит непосредственным источником реакционной способности молекул; преобразуясь снова в стационарную форму энергии химической связи, он энергетически обеспечивает видоизменение веществ, их преобразование, т. е. их обмен в организме.
Все сказанное еще раз подчеркивает тесную взаимозависимость обмена веществ и обмена энергии.
Природа макроэргических связей. Какие же закономерности в строении молекул макроэргических соединений обеспечивают их особую роль в обмене веществ и энергии? Чтобы ответить на этот вопрос, рассмотрим структуру молекулы аденозинтрифосфорной кислоты (АТФ) — наиболее важного макроэргического соединения в организме. Впервые внимание на ее роль в энергетическом обеспечении химических процессов обратил Ф. Липманн (1939—1941), а механизм распада под действием миозина и переход энергии макроэргической связи в механическую энергию сокращения мышцы исследовали В. А. Энгельгардт и М. Н. Любимова (1939).
Энергия химической связи между двумя концевыми фосфатными остатками в молекуле макроэргического соединения (АТФ) и энергия связи между остатком фосфорной кислоты и глюкозы в молекуле немакроэргического соединения (глюкозо-6-фосфата) примерно одинакова. Работу, выражающуюся одной и той же величиной, в обоих случаях необходимо затратить для отрыва фосфорного остатка и от того, и от другого соединения. Однако при изучении реакций гидролиза АТФ и глюкозо-6-фосфата выявлена существенная разница в энергетическом балансе этих одноименных реакций: в случае АТФ изменение свободной энергии составляет 32,5—34,7 кДж/моль, в случае глюкозо-6-фосфата — всего 13,1 кДж/моль.
Отсюда следует, что понятия «энергия связи» и «макроэргическая связь» совершенно различны. Первое сводится к характеристике энергетического уровня химической связи с точки зрения физической химии, т. е. как величины энергии, необходимой для разрыва связи между атомами. Второе состоит в учете энергетического эффекта в результате преобразования связи посредством химической реакции.
Здесь-то и выявляется характерная разница между обычной и макроэргической связью; последнюю нельзя рассматривать как химическую связь, обладающую повышенным по сравнению с обычной запасом энергии, или как связь непрочную, которая только и «ждет» случая, чтобы разрушиться взрывоподобно, с выделением энергии. Ее основная характеристика состоит в том, что при гидролизе или преобразовании иным путем такой связи выделяется гораздо больше энергии, чем при гидролизе или преобразовании обычной связи.
Исследование структуры той части молекулы АТФ, где сосредоточены макроэргические связи, показало, что строение ее своеобразно. В частности, распределение зарядов у атомов Р и О в трифосфатной группировке АТФ таково:
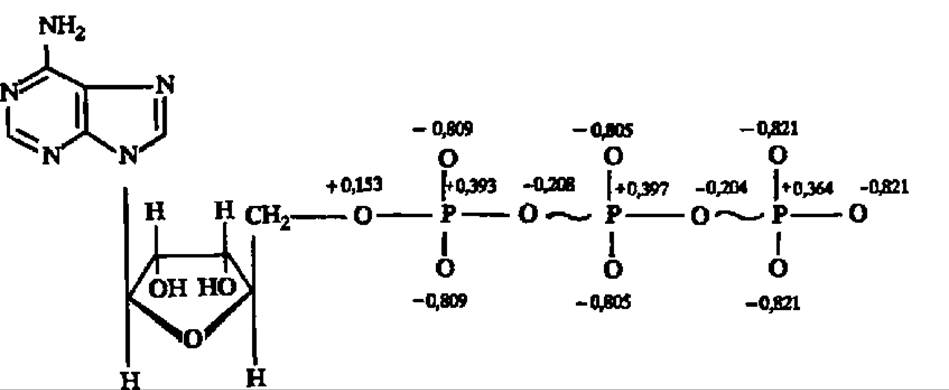
Как видно из приведенной выше формулы аденозинтрифосфорной кислоты, кислородно-фосфорный остов трифосфатной группировки составлен положительно заряженными атомами О и Р. Электростатическое отталкивание положительных зарядов в остове трифосфатной части АТФ имеет, видимо, существенное значение для выделения повышенного количества энергии при гидролитическом отрыве фосфатных групп. Аналогично, с точки зрения квантово-механических представлений, построены и остальные макроэргические соединения: для них характерно накопление положительных зарядов у атомов, непосредственно связанных макроэргической связью, и отрицательных — у окружающих эти связи атомов:
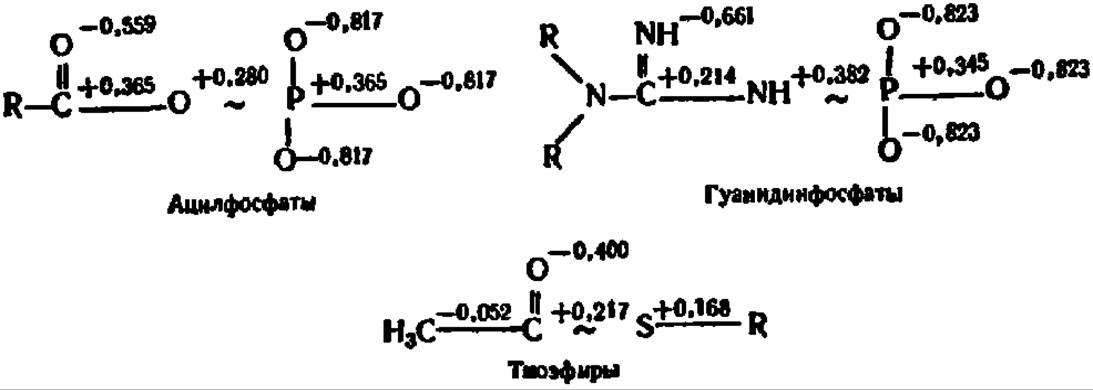
Существенно также, что п-электроны атомов О, N и S, связывающих группировки атомов в макроэргических соединениях, не в состоянии удовлетворить потребность в электронах этих группировок.
В качестве иллюстрации приведем структуру ацетилфосфата:
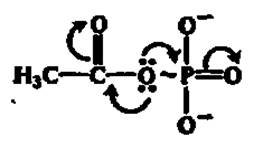
Как следствие этого, неустойчивая система сопряжения электронов в макроэргических соединениях при гидролизе и других преобразованиях легко и необратимо нарушается.
Роль макроэргических соединений в трансформации энергии. Рассмотрим теперь, как можно представить механизм превращения стабильной энергии химической связи в подвижную энергию возбуждения молекулы и обратного процесса — преобразования мобильной энергии возбуждения в стационарную энергию химической связи. Именно в этом заключается одна из наиболее существенных функций макроэргических соединений. Молекула АТФ осуществляет указанную трансформацию энергии, будучи связана со специфическим белком. Закрепление АТФ на белковой молекуле сопровождается сближением трифосфатной части молекулы с пуриновой ее частью. Выделяющаяся при распаде макроэргической связи между остатками фосфорной кислоты энергия передается на пуриновую часть молекулы. Она трансформируется при этом в мобильную энергию возбуждения электронов системы сопряженных двойных связей пуринового цикла, откуда поступает далее к месту химической реакции, переводя в возбужденное состояние электроны преобразуемого органического соединения. Это обеспечивает последнему повышенную реакционную способность, создавая, в частности, потенциальную возможность для обратного преобразования подвижной энергии возбуждения в стационарную энергию новых химических связей (рис. 63).
Энергия, которая выделяется при распаде макроэргических соединений и за счет которой может быть совершена та или иная работа, используется не только для химического синтеза. Она может служить в организме для теплообразования, свечения, накопления электричества, выполнения механической работы и т. п. При этом химическая энергия преобразуется в тепловую, лучистую, электрическую, механическую и пр. Принципиально важно то, что преобразование химической энергии в другие ее виды протекает в организме при обязательном участии соединений с макроэргическими связями, в частности АТФ. В молекуле АТФ, как было показано выше, происходит трансформация стабильной энергии макроэргических межфосфатных химических связей в подвижную энергию возбуждения электронов пуриновой части молекулы; это и есть, вероятно, первый этап преобразования энергии в организме. Именно поэтому АТФ занимает центральное место в энергетическом обмене живой материи.
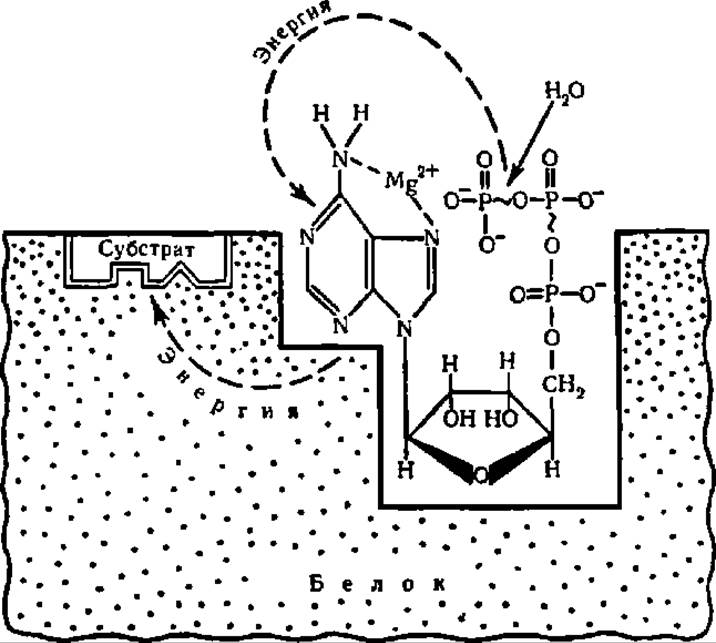
Рис. 63. Трансформация энергии в молекуле АТФ (пояснения в тексте)
Рассмотрение механизма трансформации и передачи энергии при распаде АТФ и аналогичных ей макроэргических соединений позволяет понять одно из элементарных фундаментальных свойств живой материи. Оно состоит в том, что в системах, обладающих свойствами живого, энергия, необходимая для осуществления химической реакции, будучи высвобождена в одной точке, может быть передана в другую точку, где она непосредственно используется. Это значит, что в живой природе нет необходимости в непосредственном контакте путем соударения (что является характерным свойством реакций в неживой природе) молекулы, поставляющей энергию, с молекулой, нуждающейся в энергии. Это принципиально отличает ход химического процесса в живых объектах от такового в неживых.
В живой природе идет непрерывный процесс запасания энергии в виде химической энергии связей органических веществ. Он состоит главным образом в преобразовании лучистой энергии в химическую (фотосинтез зеленых растений) и в небольшой мере — в переходе химической энергии из неорганических соединений в органические (хемосинтез микроорганизмов).
Как видно из рис. 64, АТФ играет выдающуюся роль как при запасании, так и при расходовании энергии, т. е. является ключевым веществом в энергетическом обмене организма. Известно много реакций, при посредстве которых АТФ возникает из других макроэргических соединений, и наоборот, есть много процессов, приводящих к синтезу макроэргических соединений при участии АТФ. Такие, например, макроэргические соединения, как креатинфосфат, фосфоенолпировиноградная кислота и 1,3-дифосфоглицериновая кислота, при взаимодействии с АДФ образуют АТФ с выделением креатина, пировиноградной кислоты и 3-фосфоглицериновой кислоты. Эти и подобные им соединения принято обозначать как АТФ-генерирующие вещества. Перечисленные реакции обратимы, и при известных условиях равновесие может быть смещено в сторону распада АТФ.
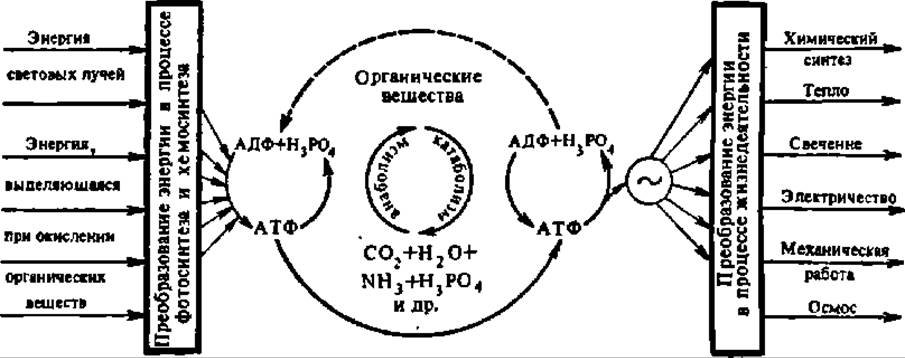
Рис. 64. Превращения энергии в живой природе (пояснение в тексте)
Общие закономерности трансформации энергии в живых системах. Обмен энергии в процессе жизнедеятельности не исчерпывается превращением химической энергии в другие виды ее и наоборот (рис. 64); он носит более широкий характер. Так, в палочках и колбочках сетчатки глаза световая энергия превращается в электрическую; в специфических структурах внутреннего уха звуковая и гидродинамическая энергия переходит в электрическую и т. п.
Трансформация одного вида энергии в другой осуществляется в организмах в морфологически разнообразных элементах — хлоропластах, мышцах (российские кардиологи недавно впервые выявили новые пути передачи энергии от места ее образования в митохондриях к точке потребления в миофибриллах сердечной мышцы), рецепторных аппаратах тканей и органов, сетчатке глаза, люминесцентных органах и т. п.
Однако всем этим разнообразным элементам свойственны некоторые общие черты строения. Они отличаются наличием двухслойных мембран с высоким содержанием липопротеинов в них и присутствием структурного белка, связывающего в упорядоченные образования достаточно унифицированные элементарные частицы. Последние включают в свой состав молекулы определенного строения, которые, собственно, и осуществляют процесс трансформации энергии. При этом энергия одного вида поглощается молекулой-преобразователем и превращается в энергию другого вида. Простейшим примером механизма внутримолекулярного превращения энергии молекулой-преобразователем служит переход стационарной энергии химических связей трифосфатной группировки молекулы АТФ в подвижную энергию возбуждения электронов ее пуриновой части (см. рис. 63). Более сложным примером являются конформационные изменения белковых молекул в процессе преобразования одного вида энергии в другой (например, мышечное сокращение).
Обмен веществ и энергии представляет единый, неразрывный процесс, где видоизменение вещества всегда сопровождается выделением или поглощением свободной энергии и где выделившаяся или поглотившаяся в том или ином количестве энергия обеспечивает распад или синтез химических связей, т. е. по существу видоизменение самих веществ. Таким образом, закономерности обмена веществ и энергии в живой природе являются частным случаем общего закона сохранения материи и энергии.