Основы биохимии - Филиппович Ю. Б. 1999
Взаимосвязь и регуляция обмена веществ
Взаимосвязь процессов обмена веществ
Было бы большой ошибкой думать, что обмен различных классов органических соединений осуществляется независимо друг от друга. Между тем в такую ошибку легко впасть, так как по необходимости изучение превращений веществ, принадлежащих к различным классам, ведется раздельно. Поэтому, рассмотрев вопросы обмена белков, нуклеиновых кислот, углеводов и липидов, необходимо разобраться во взаимосвязях между этими процессами и закономерностями их регуляции.
Если обратиться к первичному биосинтезу органического вещества, то легко убедиться в том, что первым стабильным соединением, которое образуется в результате фиксации СО2 на рибулозо-1,5-дифосфате, является 3-фосфоглицериновая кислота. Уже от этого простейшего соединения начинаются цепи реакций, ускоряемых ферментами, в результате которых синтезируются углеводы, аминокислоты, глицерин, высшие жирные кислоты, полиизопреноиды, стеролы и другие соединения. Из аминокислот, СО2 и NH3 возникают пуриновые и пиримидиновые основания. Следовательно, прямым продолжением первичной фиксации СО2 сразу являются многообразные процессы создания мономеров, из которых далее строятся биополимеры (полисахариды, белки, нуклеиновые кислоты и т. п.), разнообразные липиды и многие другие органические соединения, входящие в состав растений, животных и микробов.
Однако уже у автотрофов наряду с прямым, первичным биосинтезом органических веществ осуществляется новообразование органических соединений одних классов за счет таковых других классов. Такого рода превращения достигают своего расцвета у гетеротрофов, где не только в процессе питания, но и в ходе жизнедеятельности идет перестройка белков, нуклеиновых кислот, углеводов, липидов и многих других соединений через ключевые метаболиты промежуточного обмена, в первую очередь через пировиноградную кислоту (ПВК), а-кетоглутаровую и щавелевоуксусную кислоту (ЩУК) и ацетил-КоА (схема 14).
Следовательно, взаимопереходы между отдельными классами органических соединений — естественное, неизбежное и крупномасштабное явление в живой природе.
Рассмотрим несколько подробнее, как они осуществляются в ряде конкретных случаев.
Взаимосвязь обмена нуклеиновых кислот и белков выражается прежде всего в том, что новообразование как нуклеозидтрифосфатов, так и самих нуклеиновых кислот зависит от наличия в клеточном содержимом соответствующего набора белков-ферментов (ДНК- и РНК-полимераз, лигаз, топоизомераз, а также ферментов биосинтеза пуриновых и пиримидиновых циклов). Кроме того, именно аминокислоты (аспарагиновая — в случае пиримидиновых нуклеотидов и глицин, аспарагиновая кислота и глутамин — в случае пуриновых нуклеотидов) служат основными исходными соединениями для построения пиримидинового и пуринового колец. Вместе с тем новообразование белков по матричной схеме невозможно без участия всех видов РНК и, естественно, ДНК, на которой в качестве матрицы возникают рибонуклеиновые кислоты. Поэтому многие исследователи полагают, что в истории развития жизни на Земле биосинтез белков представлял процесс первичный, а биосинтез нуклеиновых кислот — явление вторичное, призванное в основе своей обслуживать биосинтез белка. В свою очередь, в процессе уриколиза образуется глиоксиловая кислота. Путем переаминирования она может превращаться в глицин, и, следовательно, известная часть этой аминокислоты может возникать за счет распадающихся пуриновых оснований.
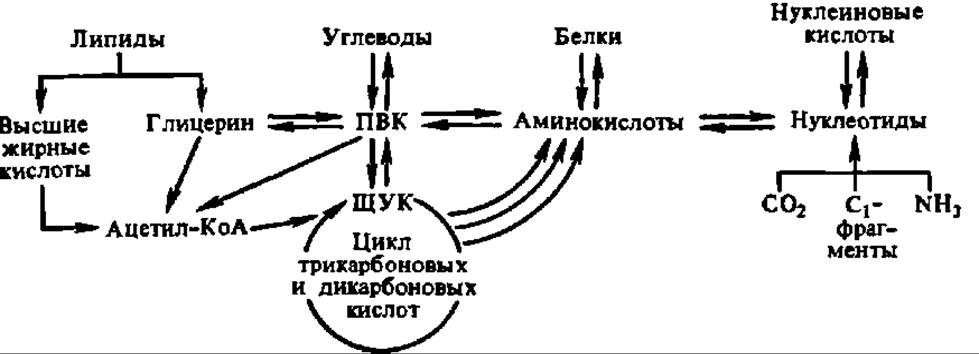
Схема 14. Взаимосвязь обмена главных классов органических соединений
Взаимосвязи в обмене нуклеиновых кислот и углеводов многообразны. Во-первых, в процессе апотомического распада углеводов образуется рибозо-5-фосфат, из которого возникает 5-фосфорибозил-1-пирофосфат, служащий совершенно незаменимым соединением для биосинтеза пуриновых и пиримидиновых нуклеотидов. Именно 5-фосфорибозил-1-пирофосфат принимает на себя недостроенную молекулу пиримидина и именно на 5-фосфорибозил-1- пирофосфате начинает строиться имидазольный цикл будущего пуринового кольца. Таким образом, ß,D-рибоза и ß,D-дезоксирибоза, являющиеся непременными составными частями пуриновых и пиримидиновых нуклеотидов, поступает в нуклеиновые кислоты за счет распадающихся углеводов.
Во-вторых, в известной мере, и распад нуклеиновых кислот может служить источником соединений, служащих для биосинтеза углеводов, так как высвобождаемая при гидролизе пуриновых и пиримидиновых нуклеотидов рибоза, включаясь в общий круговорот углеводов в организме, может переходить в рибозо-5-фосфат, из которого легко строится глюкозо-6-фосфат. Взаимопереходы последнего в фосфорные эфиры других моносахаридов широко известны.
В-третьих, распадающиеся углеводы поддерживают на определенном уровне субстратное и окислительное фосфорилирование АДФ, т. е. обеспечивают биосинтез АТФ. Последняя абсолютно необходима для превращения нуклеозидмонофосфатов в нуклеозидтрифосфаты — субстраты для полимераз. Следовательно, от наличия в организме сахаров и интенсивности их распада зависит объем биосинтеза нуклеиновых кислот. Лишь у фотосинтезирующих и хемосинтезирующих организмов эта зависимость может быть ослаблена за счет использования АТФ, возникшей в результате фотосинтетического и хемосинтетического фосфорилирования.
Наконец, в четвертых, биосинтез углеводов в значительной мере зависит от нуклеинового обмена. Эта зависимость выражается в том, что известная часть уридинтрифосфорной кислоты используется для биосинтеза УДФ-глюкозы — важнейшего продукта, гликозидные остатки с которого переносятся на нередуцирующий конец молекулы синтезируемого глюкана. Аналогична роль гуанозиндйфосфатглюкозы в биосинтезе целлюлозы и ряда других нуклеозиддифосфатсахаров в новообразовании тех или иных гексозанов и пентозанов. Все это совершенно по-новому ставит вопрос о зависимости специфического биосинтеза сложных углеводов от обмена соединений нуклеотидной природы.
Формы связи обмена нуклеиновых кислот и липидов разработаны мало. Ни те ни другие не являются непосредственными источниками соединений, которые могли бы использоваться для построения нуклеиновых кислот за счет липидов или наоборот. Следовательно, «субстратная» форма связи не характерна для обмена нуклеиновых кислот и липидов, хотя, конечно, через посредство углеводов и белков в конце концов может осуществляться частичный переход от первых ко вторым и обратно.
Что касается иного типа взаимосвязей обмена нуклеиновых кислот и липидов, то они выявляются более отчетливо. При распаде пиримидиновых оснований возникает ß-аланин — аминокислота, используемая для биосинтеза коэнзима А, столь необходимого как для новообразования, так и для деструкции высших жирных кислот. Несомненно, что ß-окисление высших жирных кислот — составных частей большинства липидов — служит источником для поддержания на достаточном уровне синтеза нуклеозид- трнфосфатов, если указанное окисление сопряжено с фосфорилированием и новообразованием АТФ. Так же, как и в биосинтезе углеводов, большую роль в биосинтезе некоторых липидов играют нуклеозиддифосфатсоединения, для образования которых расходуются соответствующие нуклеозидтрифосфаты. Так, для биосинтеза ЦДФ-холина или ЦДФ-коламина — важнейших метаболитов в синтезе фосфатидов — необходим ЦТФ — метаболит нуклеинового обмена.
Связующим звеном в обмене белков и углеводов при переходе первых во вторые и особенно вторых в первые служит ПВК. Являясь главным конечным продуктом дихотомического распада углеводов, ПВК служит исходным веществом для биосинтеза аланина, валина и лейцина. При ее карбоксилировании образуется щавелевоуксусная кислота, из которой строится новая группа аминокислот — аспарагиновая кислота, треонин, метионин, изолейцин и лизин. Вступая в цикл трикарбоновых и дикарбоновых кислот, ПВК используется для биосинтеза а-кетоглутаровой кислоты, из которой образуются глутаминовая кислота, пролин и аргинин. Предшественник ПВК — 3-фосфоглицериновая кислота — является исходным соединением для синтеза серина, глицина, цистина и цистеина.
Наконец, промежуточные продукты апотомического и дихотомического распада углеводов незаменимы в синтезе остальных постоянно встречающихся в белках аминокислот: на рибозо-5-фосфате строится имидазольное кольцо гистидина, а из эритрозо-4-фосфата и фосфоенолпировиноградной кислоты синтезируется шикимовая кислота, из которой образуются фенилаланин, тирозин и триптофан. Таким образом, у аутотрофов из углеводов при наличии источника аммиака в организме могут синтезироваться все аминокислоты, постоянно встречающиеся в белках. Естественно, что из них образуются белки, и, следовательно, переход углеводов в белковые тела представляет основной вид взаимосвязи обмена указанных двух классов соединений.
Возможен и обратный процесс. Многие аминокислоты (аланин, фенилаланин, тирозин, гистидин, триптофан, серин, цистеин) содержат в своем составе трехуглеродный фрагмент, из которого в процессе распада указанных аминокислот возникают ПВК и ее дериваты. Дезаминирование глутаминовой и аспарагиновой кислот ведет к образованию а-кетоглутаровой и щавелевоуксусной кислот соответственно, которые при посредстве цикла трикарбоновых и дикарбоновых кислот переходят в ПВК. Такова же судьба пролина, который легко превращается в глутаминовую кислоту, а из нее — в пировиноградную. Следовательно, подавляющее большинство аминокислот может явиться в организме источником для образования ПВК. От последней несложен переход к углеводам посредством в основном обращения реакций дихотомического распада фруктозо-1,6-дифосфата.
Из других форм взаимосвязи обмена белков и углеводов привлекают внимание две. Многочисленные белки-ферменты обслуживают процессы распада и синтеза углеводов в организме. В свою очередь, распад углеводов, сопряженный с синтезом АТФ из АДФ и неорганического фосфата, энергетически обеспечивает белковый синтез в клетке.
Взаимосвязь обмена белков и липидов выражается в том, что распад липидов, как и распад углеводов, обеспечивает, с одной стороны, исходные соединения для биосинтеза аминокислот (а из них белков) и, с другой стороны, не менее, а может быть, более, чем углеводы, поддерживает образование белков энергетически.
Одним из основных продуктов распада липидов, в частности высших жирных кислот, возникающих при гидролизе триглицеридов, фосфатидов или стеридов, является ацетил-КоА. Включаясь в цикл трикарбоновых и дикарбоновых кислот, он обеспечивает синтез а-кетоглутаровой кислоты, превращение которой в аминокислоты рассмотрено выше. Поступая в глиоксилевый цикл, ацетил-КоА служит для расширенного воспроизводства в организме щавелевоуксусной кислоты, а из нее — ПВК. Из обеих названных кислот также синтезируются аминокислоты.
Обмен глицерина, высвобождаемого при гидролизе триглицеридов, через углеводы ведет к таким аминокислотам, как гистидин, фенилаланин, тирозин и триптофан. Следовательно, все постоянно встречающиеся в белках аминокислоты могут синтезироваться за счет распадающихся липидов.
В известной мере, возможен синтез липидов за счет распадающихся белков. В предыдущем разделе было показано, что при распаде ряда аминокислот образуется ПВК. При ее окислительном декарбоксилировании возникает ацетил-КоА — исходное соединение для синтеза высших жирных кислот, стеролов и других составных частей липидов. ПВК может также превратиться в фосфоглицерин (путем обращения реакций дихотомического распада углеводов) — другой важный компонент липидов. Однако такого рода переход вряд ли широко осуществляется в нормальных условиях жизнедеятельности.
Энергетическая роль липидоц, особенно триглицеридов, общеизвестна. Потенциальные возможности для синтеза АТФ сопряженно с окислением высших жирных кислот огромны. Известны случаи, когда распад липидов является единственным источником энергии для биосинтеза белка (например, при синтезе фиброина и серицина шелка в шелкоотделительной железе коконопрядущих насекомых).
Говоря о взаимосвязи обмена белков и липидов, нельзя обойти вопрос о влиянии последних на процесс биосинтеза белков. Твердо установлено, что рибосомальный синтез белка протекает во много раз энергичнее, если рибосомы связаны с липопротеиновыми мембранами.
Углеводы и липиды очень легко взаимопревращаготся в организме; связующими соединениями при этих переходах служат ПВК в ацетил-КоА.
Пировиноградная кислота — основной продукт дихотомического распада углеводов, при окислительном декарбоксилировании дает ацетил-КоА, который служит для синтеза высших жирных кислот, стеролов, каротиноидов и других полиизопреноидов. Столь же легко осуществляется переход от углеводов к фосфоглицерину, необходимому для синтеза простых и сложных липидов.
Ацетил-КоА и глицерин — главные продукты распада липидов — служат исходными соединениями для синтеза углеводов. Ацетил-КоА при посредстве глиоксилевого цикла переходит в ПВК, а из нее — в углеводы путем обращения реакций дихотомического распада последних.
Превращение глицерина в углеводы идет через 3-фосфоглицериновый альдегид, а затем описанным выше способом.
Сказанное не исчерпывает всего многообразия взаимосвязей обмена белков, нуклеиновых кислот, углеводов, липидов и других соединений. Между ними существуют более сложные, нежели простое использование в качестве субстратов, формы взаимозависимости. Так, вещества, образующиеся в процессе обмена соединений одного класса, оказывают глубочайшее влияние на обмен веществ, относящихся к другому классу. Превосходным примером подобного типа взаимосвязи обмена белков и нуклеиновых кислот может служить образование информационной РНК, служащей матрицей для биосинтеза специфических белков, с одной стороны, и блокирование синтеза иРНК определенного вида белками — с другой.
Общеизвестно, что никакие реакции обмена невозможны без специфических белков-ферментов, и в этом смысле белковый обмен определяет ход превращений соединений, относящихся к другим классам. Решающее значение имеет ход окислительного фосфорилирования и создание резервов АТФ в клетке. От уровня последней в клеточном содержимом зависит, в свою очередь, весь ход обмена веществ, ибо АТФ обеспечивает энергетические потребности биосинтеза соединений всех классов. Число подобных примеров глобальной взаимозависимости и взаимообусловленности обмена белков, нуклеиновых кислот, углеводов, липидов и других соединений огромно. В совокупности они и составляют учение о регуляции обмена веществ. Но каждый из них в отдельности подчеркивает ту или иную форму взаимосвязи обмена веществ в организме.