Химия и биология белков - Ф. Гауровитц 1953
Синтез белка
Механизм синтеза белков
В предыдущих разделах данной главы были изложены немногочисленные имеющиеся данные относительно синтеза белков путем полимеризации и ферментативной конденсации. В некоторой степени был также освещен вопрос о месте синтеза белков в клетках и о связи этого процесса с другими процессами обмена. Результаты проведенных до настоящего времени исследований могут быть суммированы в следующих положениях.
1. Синтез белков происходит, повидимому, в ядрышке или около него, а также в малых гранулах цитоплазмы клеток и в частицах вирусов.
2. Все частицы, в которых происходит синтез белков, содержат много рибонуклеиновой кислоты.
Этих данных, конечно, совершенно недостаточно для построения более или менее обоснованной теории механизма синтеза белков. Самое большее, что мы в настоящее время в состоянии сделать, это высказать только некоторые предположения по этому вопросу и рассмотреть, в какой степени эти предположения соответствуют современным данным о физико-химических свойствах белков. Основным вопросом всей проблемы синтеза белков является вопрос о том, каким образом в организме образуются белки, обладающие высокой специфичностью. В предыдущих главах данной книги неоднократно подчеркивалось, что каждый вид животных имеет свои специфичные белки и что белки многих органов тоже обладают определенной специфичностью, которая отличает их от белков других органов того же животного. Специфичность белков определяется их аминокислотным составом, порядком расположения аминокислот в пептидной цепи и специфической формой скрученных пептидных цепей.
Каким же образом можно объяснить все эти факты? Многие исследователи полагают, что в клетке содержится некая неизменяющаяся структура, нечто вроде шаблона или матрицы, при помощи которой происходит синтез белка. Хотя никто никогда не наблюдал этого шаблона и нет никаких доказательств его существования, тем не менее мы все же вынуждены принять это объяснение, так как у нас нет в настоящее время иных способов объяснить постоянное образование одних и тех же специфических белков в каждой клетке.
В связи с основным вопросом о происхождении специфичности белков возникают следующие вопросы: 1) Идентична ли форма образующегося в клетке белка форме шаблона или же она представляет собой негативный отпечаток этого шаблона? 2) Состоит ли шаблон из белков, нуклеиновых кислот, нуклеопротеидов или же он построен из других соединений?
Различные авторы дают разноречивые ответы на эти вопросы. Некоторые из них считают, что синтез белков представляет собой в основном репродукцию, т. е. что синтезированные белки образуют позитивный отпечаток шаблона; другие же, наоборот, полагают, что синтезированные белки представляют собой его негативный отпечаток. Нет также единомыслия и по вопросу о роли нуклеиновых кислот в синтезе белков.
Гипотезы можно классифицировать следующим образом [62].
1. Нуклеиновые кислоты образуют шаблон, по которому формируются синтезирующиеся белки.
2. Нуклеиновые кислоты реагируют с белками, образуя нерастворимые соединения, вследствие чего синтезируемые белки постоянно удаляются из раствора. Благодаря этому равновесие между предшественниками белков и синтезируемыми белками все время сдвинуто вправо, что создает условия для дальнейшего синтеза белков.
3. Нуклеиновые кислоты поставляют энергию для синтеза белков; она может быть получена при окислении пентозы или пуриновых оснований, входящих в состав нуклеиновых кислот.
4. Нуклеиновые кислоты являются материалом для образования коферментов, так как рибоза, аденин и фосфорная кислота могут быть использованы для образования флавиновых и пиридиновых нуклеотидов.
При рассмотрении изложенных выше взглядов необходимо прежде всего обсудить вопрос о том, представляют ли собой молекулы белка позитивные или негативные отпечатки гипотетического клеточного шаблона. Мы знаем, что молекулы белка состоят из длинных пептидных цепей и что эти цепи сложены в складки таким образом, что образуется трехмерная глобулярная молекула. Диаметр этой глобулярной частицы может колебаться от 20 до 100 А. В настоящее время мы не знаем таких сил, при помощи которых указанный шаблон мог бы действовать на столь большом расстоянии. Действие сил, обусловливающих реакцию между ионами и полярными группами (см. гл. X), резко уменьшается по мере увеличения расстояния между этими группами, и действие их сказывается лишь на расстоянии, не превышающем 4—5 Å. Лишь в том случае, если бы указанные реагирующие группы находились в вакууме, эти силы могли бы действовать на расстоянии больше 4—5 А. Однако в биологических системах мы имеем дело не с вакуумом, а с системами, в которых между реагирующими группами всегда находятся молекулы воды, представляющие собой полярные частицы с высокой диэлектрической постоянной.
Действие сил, обусловливающих электростатическое притяжение и отталкивание, будет поэтому значительно уменьшено противодействием молекул воды и не будет превышать 1/80 того действия, которое эти силы оказывают в вакууме. Иными словами, действие электростатических сил в водных растворах распространяется на расстояние не свыше 4—5 Å. Из этих соображений ясно, что предположение о прямой репродукции глобулярной молекулы диаметром 20—100 Å при помощи межмолекулярных сил несовместимо с имеющимися данными о дальности действия и эффективности этих сил.
С этими данными [135—137] согласуется одна-единственная гипотеза, высказанная много лет назад и состоящая в том, что репродукция молекул белка может происходить только в том случае, если соответствующие реакции протекают в растянутых мономолекулярных пленках [133, 134]. Согласно этой гипотезе, образование пептидных цепей молекулы белка происходит в мономолекулярной пленке, растянутой на поверхности шаблона; образование же складок и превращение двумерного-пептидного слоя в трехмерную глобулу белка представляет собой вторичный процесс, происходящий в другом месте и отличающийся от процесса истинного синтеза пептидных цепей.
Ряд физико-химических соображений заставляет автора данной книги принять, что вышеуказанная двумерная пептидная структура представляет собой позитивный отпечаток шаблона.
Это положение на первый взгляд противоречит представлению о том, что форма молекулы антитела геометрически дополняет форму молекулы антигена, а форма молекулы фермента дополняет форму молекулы субстрата. В гл. XIV, рассматривая соответствие между формой молекул антигена и антитела, мы указывали, что антиген в этом случае является шаблоном, а молекулы антитела его негативными отпечатками. Этот взгляд, подтверждаемый всеми имеющимися в настоящее время данными об образовании антител и реакции антиген—антитело, не противоречит, однако, предположению об образовании пептидных цепей белка в мономолекулярном слое. Легко можно допустить, что антиген играет роль шаблона в упомянутой вторичной реакции, а именно: в процессе превращения растянутой двумерной пептидной пленки в трехмерную глобулярную молекулу антитела. В соответствии с этим форма молекулы антитела геометрически дополняет форму глобулярной молекулы антигена, а не форму его растянутой молекулярной пленки. Синтез антител представляет собой частный случай синтеза белков, и нет никаких оснований думать, что он чем-либо существенным отличается от обычного синтеза других белков, например сывороточных глобулинов, т. е. что существуют два различных механизма синтеза белков. Можно поэтому считать, что и синтез белков, протекающий в нормальных клетках, также приводит к образованию молекул, форма которых геометрически дополняет форму детерминирующих групп находящихся рядом молекул шаблона. Таким образом, мы обобщаем представление о геометрически дополняющих друг друга формах, распространяя его не только на синтез антител, но и на синтез всех других белков. Обычные клеточные белки, образующиеся в норме, было предложено называть аутоантителами [138], чтобы обозначить этим, что форма их молекул геометрически дополняет форму молекул шаблонов, находящихся в месте их образования. Только в том случае, когда нормальные шаблоны покрываются или обволакиваются чужеродным белком антигена, естественный процесс превращения растянутых пептидных цепей в обычные глобулярные молекулы нарушается и взамен его происходит синтез антител с молекулами, геометрически дополняющими по своей форме молекулы антигена [135].
Представление о шаблоне, на котором образуется негативный отпечаток, было использовано многими авторами для объяснения механизма образования различных белков [139, 140]. В частности, образование белков раковой ткани рассматривалось как следствие того, что в этой ткани шаблоны имеют ненормальное строение [142], обусловленное либо лежащими в одной плоскости кольцами канцерогенных веществ [141], либо их электронной структурой [143].
Автор полностью разделяет мнение Паулинга [144] о том, что все специфические биологические процессы обусловлены одними и теми же межмолекулярными силами, действие которых, в свою очередь, определяется контактом атомов и процессом образования отпечатков. Развивая эту концепцию, автор постулирует, что первой ступенью синтеза белка является образование позитивного отпечатка с первичного шаблона, причем и шаблон и отпечаток представляют собой мономолекулярные пленки. Второй же ступенью является образование негативного глобулярного отпечатка, форма которого геометрически дополняет форму второго, также глобулярного шаблона.
Если мы примем эту концепцию для процессов синтеза белков в клетках in vivo, то необходимо ответить и на ряд вопросов, касающихся природы шаблона и механизма его действия. Каким образом в действительности на этом двумерном шаблоне образуется растянутая пленка белка? Представляет ли собой шаблон белок, нуклеиновую кислоту или какое-либо другое соединение?
Как известно, нуклеиновые кислоты всегда присутствуют в тех местах клеток, где происходит синтез белков, это дало многим авторам основание считать, что именно нуклеиновые кислоты образуют предполагаемые шаблоны для синтеза белков и что именно они обусловливают специфичность всех вновь образуемых белков в организме животных и растений.
Нуклеиновые кислоты являются полярными соединениями, обладающими многими отрицательно заряженными группами фосфорной кислоты, полярными гидрофильными группами гидроксилов сахаров и положительно заряженными группировками, входящими в состав пуриновых и пиримидиновых оснований. Нет сомнения в том, что все эти группировки могут оказывать большое влияние на соседние молекулы. Выше (см. гл. XI) уже приводились убедительные данные о том, что при соединении нуклеиновых кислот с белками изменяется форма молекул как белка, так и нуклеиновых кислот [145].
Предполагают, что положительно заряженные группы диаминокислот белка реагируют с отрицательно заряженными группами фосфорной кислоты нуклеиновых кислот и что таким образом возникает негативный отпечаток нуклеиновой кислоты, которая выполняет в данном случае функцию шаблона [140, 146]. Однако, по мнению автора, высокая кислотность нуклеиновых кислот вряд ли может служить доказательством того, что именно они играют роль специфического шаблона, так как их кислотность настолько велика, что они неспецифически реагируют со многими белками. Кроме того, необходимо принять во внимание, что нуклеиновые кислоты состоят только из семи или восьми различных составных частей: аденина, гуанина, цитозина, урацила, тимина, фосфорной кислоты и рибозы или дезоксирибозы. Трудно поэтому представить себе, что нуклеиновая кислота как шаблон может определить столь небольшие различия между белками, как те, которые наблюдаются, например, между сывороточными альбуминами человека и быка, лишь незначительно отличающимися друг от друга по своему аминокислотному составу [147]. Далее, мы до сих пор не знаем, обладают ли нуклеиновые кислоты видовой специфичностью. Если они не обладают этим свойством, то они вообще не могут образовывать специфических шаблонов. Однако если даже допустить видовую специфичность нуклеиновых кислот, то очень трудно представить себе, каким образом могут они определять специфичное положение различных аминокислот в образующейся пептидной цепи, поскольку фосфорная кислота нуклеиновых кислот должна реагировать главным образом с щелочными аминокислотами белка.
Остается до сих пор неизвестным, имеется ли какая-либо последовательность в чередовании пуриновых оснований в молекуле нуклеиновых кислот. Мы знаем лишь, что они состоят из мононуклеотидов, из которых каждый содержит одну группу фосфорной кислоты. Расстояния между этими группами в растянутой полинуклеотидной молекуле одинаковы. Отрицательно заряженные группы фосфорной кислоты отталкивают друг друга, вследствие чего вся молекула имеет тенденцию оставаться в растянутом положении.
В настоящее время нет никакой возможности дать ответ на вопрос о том, каким образом эта более или менее правильно построенная молекула нуклеиновой кислоты может определять образование бесконечного числа специфических белков. В связи с этим совершенно нецелесообразно обсуждать далее гипотезу, приписывающую нуклеиновой кислоте роль первичного шаблона, по которому формируются мономолекулярные белковые пленки. Мы перейдем поэтому к рассмотрению тех взглядов, согласно которым этот первичный шаблон образован слоем белка.
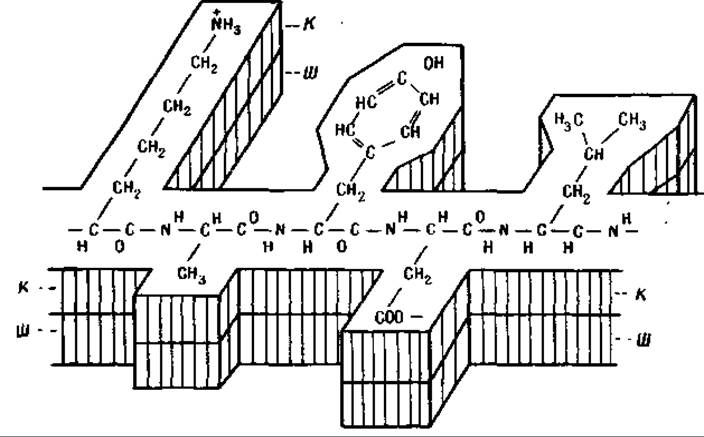
Фиг. 51. Дублирование белкового шаблона [135]. Пептидные цепи шаблона и копии образованы лизином, аланином, тирозином, аспарагиновой кислотой и лейцином.
Ш — шаблон; К — копия.
По этой гипотезе вновь образованная растянутая белковая пленка должна представлять собой копию другой растянутой белковой пленки [135, 136]. Мы должны вообразить в этом случае, что каждая из l-аминокислот, находящихся в слое шаблона, притягивает из окружающей среды того же типа l-аминокислоту и что аминокислоты, входящие в образующуюся копию, связываются друг с другом благодаря действию неспецифических протеолитических ферментов типа трипсина или папаина. В этом случае образование копии может произойти так, как это схематически представлено на фиг. 51, где пептидные цепи шаблона и его копии образованы лизином, аланином, тирозином, аспарагиновой кислотой и лейцином.
Изложенное выше представление, согласно которому определяющим моментом всего процесса синтеза белков является специфическая адсорбция аминокислотами шаблона тех же аминокислот из окружающей среды, довольно хорошо согласуется с рядом прочно установившихся химических и физикохимических воззрений.
Этот процесс адсорбции можно сравнить с процессом кристаллизации, который, как известно, также отличается высокой специфичностью. Если мы насытим раствор, содержащий много различных аминокислот, в отношении одной из них так, что часть аминокислоты останется в осадке, и затем будем медленно выпаривать раствор, то только молекулы этой аминокислоты будут адсорбироваться на кристаллических частицах нерастворенного осадка, в связи с чем эти частицы будут увеличиваться в размерах. Никогда еще не удавалось наблюдать, чтобы какая- либо другая аминокислота включалась в решетку растущего кристалла. Если мы выпарим раствор до такой степени, что начнут осаждаться другие аминокислоты, то они будут образовывать в большинстве случаев отдельные кристаллы. Из этого примера видно, что адсорбция находящихся в растворе молекул решетками тех же молекул, находящихся в осадке, представляет собой в высшей степени специфический процесс.
Указанная аналогия между специфичностью кристаллизации и биологической специфичностью привлекла внимание многих биологов и химиков [133, 135, 144, 148]. Идея о том, что твердая частица какого-либо вещества может служить ядром, на которое накладываются другие частицы этого вещества, получила известное подтверждение в ряде открытий в других областях биохимии. Одним из таких открытий является установление роли, которую играет каркас гликогена в ферментативном синтезе новых молекул гликогена.
Работами Кори и Кори [149] было установлено, что природа полисахарида, образующегося при ферментативном синтезе полисахаридов из глюкозо-1-фосфата, зависит от того, следы какого полисахарида присутствуют в растворе. Если в качестве затравки прибавлен полисахарид, имеющий разветвленные цепи, то образуются полисахариды с разветвленными цепями. Если же прибавлен полисахарид с неразветвленными цепями, то в результате ферментативного синтеза будут образовываться только такие полисахариды. Прибавленный полисахарид можно, таким образом, рассматривать как вещество, пускающее в ход реакцию и определяющее ее направление, т. е. как затравку.
Это новое представление, несомненно, имеет очень большое значение для развития проблемы образования макромолекул в организме. Согласно этому представлению, мы должны считать, что концевые группы разветвленной или неразветвленной молекулы затравочного вещества играют роль специфического организатора, влияющего на действие неспецифического фермента.
Таким образом, отпадает необходимость допускать наличие в организме множества специфических катализаторов. Допущение же это всегда казалось маловероятным. Специфичность синтетических процессов можно теперь отнести не за счет специфичности катализаторов, а за счет специфичности организаторов, или затравок.
Эти организаторы можно представить себе в виде соединений, не обладающих ферментной активностью и аналогичных элементарным структурным единицам кристаллов, единственным назначением которых является определение формы отлагающихся на них веществ. Факты, подобные найденным Кори и Кори при изучении синтеза полисахаридов, были обнаружены и при исследовании синтеза белков.
В гл. XI уже указывалось, что гемоцианин — медьсодержащий дыхательный пигмент беспозвоночных — может быть расщеплен на частицы, молекулярный вес которых составляет лишь 1/2 или даже 1/8 молекулярного веса гемоцианина. Это расщепление гемоцианина наступает, если он находится в щелочном растворе при pH 8,5. Если же pH раствора сдвинуть в кислую сторону и довести его до 6,85, то наступает обратный процесс, а именно: ассоциация продуктов распада, и образуется первоначальная молекула гемоцианина. При этом, однако, оказалось, что фрагменты гемоцианина одного вида могут ассоциироваться только с фрагментами того же гемоцианина, но не с фрагментами гемоцианина, полученного от другого вида. Так, например, фрагменты гемоцианина Helix pomatia не ассоциируются с фрагментами гемоцианина, полученного из организма Litiorina littorea [150].
Все эти наблюдения над специфичностью роста кристаллов, ролью затравки и ассоциацией фрагментов свидетельствуют о том, что при известных условиях молекулы одного типа избирательно притягивают молекулы того же типа. Силы, обусловливающие взаимное притяжение этих молекул, во многих случаях являются чисто ионными силами.
Наиболее простым примером притяжения этого рода является рост кристалла хлористого натрия, который в основном обусловлен взаимным притяжением положительно заряженных ионов натрия и отрицательно заряженных ионов хлора. Это объяснение, однако, неприменимо к росту кристаллов неэлектролитов, например кристаллов нафталина. Хотя нафталин не содержит ни ионных, ни полярных гидроксильных или кетогрупп, все же только молекулы нафталина присоединяются к кристаллам нафталина в растворе, содержащем, кроме нафталина, и другие близкие к нему углеводороды. Специфичность роста кристаллов нафталина может быть объяснена только тем, что форма молекул нафталина, находящихся в растворе, соответствует форме молекул, образующих решетку кристаллов нафталина, вследствие чего они легче других молекул могут проникать в бреши между частями решетки. Этот процесс, таким образом, представляет собой чисто топохимический феномен, связанный с поверхностными явлениями на твердой фазе. На поверхности этой фазы могут адсорбироваться различные вещества, но всегда преимущество в этом отношении будут иметь молекулы того же типа.
Тот факт, что сродство между молекулами одного и того же типа выше, чем сродство между молекулами разных типов, хорошо известен физико-химикам.
Величина энергии связи может быть определена путем измерения теплоты растворения или теплоты плавления. Точка плавления чистых веществ всегда выше, чем точка плавления веществ, содержащих примеси; другими словами, для разделения молекул одного и того же типа требуется больше энергии, чем для разделения молекул различных типов.
Притяжение молекул будет, конечно, еще выше, если оно обусловлено взаимодействием противоположно заряженных ионных групп этих молекул. По этой причине кристаллы ионного типа тверды и имеют высокую точку плавления, тогда как кристаллы неполярного типа мягки и плавятся при низких температурах.
Изучение явления кристаллизации помогает понять процесс образования белков, происходящий путем наложения аминокислот из раствора на аминокислоты растянутой пленки шаблона. Можно, правда, возразить на это, что в пленке белка, служащего шаблоном, содержатся только остатки аминокислот, связанные друг с другом в пептидных цепях и отличающиеся по своей форме от свободных аминокислот, находящихся в растворе. Это возражение не лишено основания. Однако следует учесть, что свободные аминокислоты и аминокислотные остатки в пептидной цепи идентичны по форме своих основных частей, как видно, например, из следующих формул:
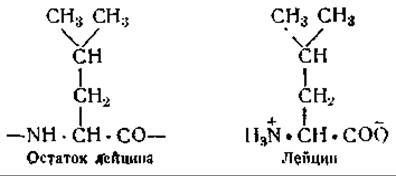
Лейцин, растворенный в клеточной жидкости, несомненно ближе подходит по своей форме к остатку лейцина в пептидной цепи, чем любая другая аминокислота (см. фиг. 51).
Согласно изложенным выше представлениям, аминокислоты, отложившиеся на поверхности шаблона, образуют затем пептидную цепь. Этот процесс является ферментативным, однако нет никаких доказательств того, что в нем принимают участие специфические ферменты, и, следовательно, нет необходимости постулировать наличие таких специфических ферментов. Протеолитические ферменты, выделенные из органов, не являются специфическими, так как они катализируют гидролиз самых различных белков животного и растительного происхождения. Эти же ферменты могут катализировать и процесс синтеза пептидов из аминокислот, что было убедительно показано Бергманом и его сотрудниками [18, 20]. В предыдущих разделах данной главы уже указывалось, что синтез белков нельзя рассматривать просто как процесс, обратный их расщеплению, и что промежуточные реакции синтеза могут протекать иначе, чем соответствующие гидролитические реакции. Наиболее важным моментом является то, что мы не имеем решительно никаких доказательств специфичности ферментов, участвующих как в гидролизе, так и в синтезе белка. Специфичность образующегося белка можно вполне удовлетворительно объяснить специфической адсорбцией аминокислот на поверхности шаблона.
Прежде чем перейти к рассмотрению вопросов, касающихся второй фазы белкового синтеза — фазы, в которой происходит свертывание пептидных цепей, — необходимо попытаться ответить на следующие два вопроса: 1) Какова роль нуклеиновых кислот? 2) Чем отличается белковая пленка шаблона от образующейся на ней копии белка? Почему только последняя образует глобулярную молекулу, а пленка шаблона остается растянутой и нерастворимой?
Повидимому, ответ на оба эти вопроса заключается в следующем.
Роль нуклеиновых кислот сводится, вероятно, к тому, чтобы удерживать белковую пленку шаблона в растянутом состоянии. В таком состоянии белковые пленки могут оставаться только под влиянием сил, действующих между поверхностями раздела. Эти силы вызывают развертывание пептидных цепей и удерживают их развернутыми в растянутой мономолекулярной пленке. Можно предположить, и это является весьма вероятным, что различные типы нуклеиновых кислот играют роль носителей белковых пленок шаблона и что замена одной нуклеиновой кислоты другой может до некоторой степени повлиять на образование копии. Это влияние не может, однако, проявиться особенно сильно, если только при этом не будет повреждена сама белковая пленка шаблона.
Переходя к рассмотрению вопроса о том, почему образующаяся копия белковой пленки свертывается с образованием растворимых глобул, тогда как белковая пленка шаблона остается нерастворимой, необходимо принять во внимание cледующее. Каждый пептидный слой имеет толщину 8—10 Å, в связи с чем глобулярная частица может состоять только из немногих пептидных слоев. Мы не знаем до сих пор, действительно ли эти пептидные слои образуют складки или же несколько двумерных слоев сочетаются друг с другом таким образом, что образуется глобулярная частица. Этот вопрос был до настоящего времени исследован только в отношении гемоглобина (см. гл. XI). Предполагаемая структура молекулы гемоглобина изображена на фиг. 52 [151]. Молекула состоит из четырех пептидных слоев, из которых каждый образован зигзагообразно свернутой пептидной цепью, уложенной в пять складок.
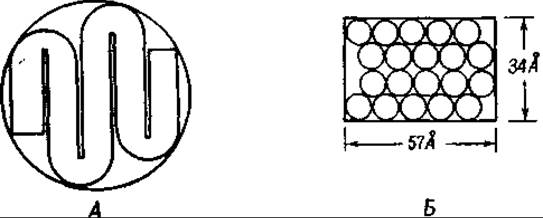
Фиг. 52. Молекула гемоглобина [151].
А — поперечный разрез, показывающий складчатые пептидные цепи; Б — вертикальный разрез, показывающий упаковку цепей.
Как уже неоднократно указывалось, результаты иммунологических исследований заставляют прийти к заключению, что трехмерные молекулы антител имеют форму, геометрически дополняющую форму молекул введенного антигена.
Антиген, очевидно, участвует в процессе скручивания пептидных цепей антитела, в результате чего образующаяся молекула антитела представляет собой негативный отпечаток детерминирующих групп антигена. При синтезе обычных белков, т. е. при отсутствии чужеродного антигена, его роль выполняют полярные молекулы, находящиеся в клетке, в связи с чем образующиеся белки можно рассматривать как аутоантитела [138]. Форма их молекул геометрически дополняет форму каких-то детерминирующих структур нормальной клетки. Роль детерминирующих структур могут играть липиды, углеводы, белки и нуклеиновые кислоты. Работами Ландштейнера было достаточно убедительно доказано, что практически все полярные соединения в той или иной степени влияют на образование антител, причем эта роль может принадлежать и таким синтетическим продуктам, как азо- и нитросоединения. Таким образом, нет оснований исключать какие-либо полярные соединения из группы веществ, влияющих на синтез белков.
В тех случаях, когда мы имеем дело с организмом высших животных или растений, мы не можем сказать ничего определенного о химической природе трехмерных шаблонов, принимающих участие во второй фазе синтеза белков, т. е. в превращении двумерной белковой пленки в трехмерную молекулу белка.
У простых вирусов жиры и углеводы практически отсутствуют. Тем не менее, хотя вирусы содержат только белки и нуклеиновые кислоты, синтез белков протекает у них очень интенсивно. Из сказанного можно заключить, что у вирусов, а возможно, и у более сложных организмов роль вторичных шаблонов выполняют главным образом белки, нуклеиновые кислоты или нуклеопротеиды.
При исследовании процессов синтеза белка у вирусов мы имеем дело еще с одним упрощением условий, так как в состав вирусов входит только один тип белка. Приходится поэтому считать, что этот белок выполняет как роль двумерного шаблона в первой фазе синтеза, так и роль трехмерного шаблона во второй фазе, если только какую-либо из этих функций не принимают на себя белки хозяина. Можно допустить, что одна часть вирусного белка, соединенная с нуклеиновой кислотой и образующая двумерный шаблон, принимает участие в первой фазе синтеза, тогда как другая часть того же белка, присутствующая в трехмерной глобулярной форме, определяет специфичность процесса во второй фазе, т. е. превращение двумерной пленки в трехмерную глобулу.
Является ли шаблон катализатором? Можно ли рассматривать введенный антиген как катализатор? Многие биохимики отвечают на эти вопросы утвердительно. Однако, по мнению автора, необходимо отличать вещества, изменяющие течение реакции, но не влияющие на ее скорость, от истинных катализаторов. Различие между этими двумя типами веществ становится очевидным, если учесть данные исследований Бредига, которые были уже детально рассмотрены Севагом [152].
Бредиг показал, что если реакция бензальдегида с синильной кислотой катализируется дизгиламином, соединенным с целлюлозой, то образуется оптически активный нитрил миндальной кислоты. Образование оптически активного изомера обусловлено, очевидно, асимметричной структурой целлюлозы и не зависит от амина, щелочные группы которого проявляют каталитическое действие и в отсутствие целлюлозы. Целлюлоза в этой реакции, очевидно, не может рассматриваться как катализатор. Для обозначения веществ, которые изменяют течение реакции таким образом, что образуется преимущественно один из возможных оптических изомеров, было предложено название «столбовые вещества» [154]. Волокна целлюлозы образуют нечто вроде столба, с которым постоянно сталкиваются молекулы бензальдегида и синильной кислоты. При этих столкновениях они ориентируются асимметричными волокнами целлюлозы таким образом, что облегчается образование одного из стереоизомеров нитрила миндальной кислоты. Целлюлозу в этой реакции можно назвать также организатором или индуктором асимметрического синтеза, чтобы подчеркнуть различие между ней и истинным катализатором — амином. Для обозначения функции целлюлозы в этой реакции более других подходит термин «организатор», так как этот термин указывает, что главной функцией целлюлозы является «организация» реакции таким образом, чтобы она протекала в строго определенном направлении. По аналогии с рассмотренным явлением следует сказать, что и гипотетические шаблоны в изложенной выше теории белкового синтеза не являются катализаторами; их роль заключается лишь в том, чтобы определенным образом адсорбировать аминокислоты, создавая тем самым условия для ферментативного образования пептидных цепей только одной определенной структуры. Эти шаблоны предотвращают, следовательно, возможность образования пептидных цепей с произвольно расположенными аминокислотами.
Суммируя все, что нам известно о механизме белкового синтеза, можно разделить этот процесс на следующие фазы:
1. Специфическая адсорбция аминокислот из клеточного содержимого идентичными аминокислотными остатками, входящими в состав растянутой белковой пленки.
2. Действие неспецифического фермента на адсорбированные аминокислоты, в результате чего образуется копия белковой пленки, служащей шаблоном.
3. Свертывание двумерной белковой пленки и образование трехмерной глобулярной молекулы. Форма этой трехмерной молекулы определяется формой и электростатическим полем полярных групп, входящих в состав клеточного содержимого и находящихся в непосредственной близости от места, где происходит синтез. В том случае, когда происходит синтез антител, форма глобулярной молекулы определяется формой детерминирующих групп антигена.
Один из основных пунктов изложенной выше гипотезы составляет представление о том, что действие ферментов не является специфическим и что специфичность синтеза белков определяется только электростатическим полем и формой поверхности шаблона. Это утверждение основано на том, что до сих пор не найдено ни одного фермента, который обнаруживал бы видовую специфичность. Так, например, при действии пепсина цыпленка на пепсиноген свиньи образуется пепсин свиньи, а не цыпленка [156]. Белки растений и животных одинаково легко расщепляются протеолитическими ферментами растительного и животного происхождения.
Несмотря на эти данные, некоторые авторы все же признают наличие ферментов, катализирующих синтез специфических белков. Так, например, Барнет [155] на основании ряда соображений, изложенных в гл. XIV (см. стр. 351), полагает, что антитела образуются не только в присутствии сенсибилизирующих антигенов, но и тогда, когда антиген разрушается или удаляется из сенсибилизированного организма. Образование антител, по мнению Барнета, свойственно не только непосредственно стимулированным клеткам, но также и тем клеткам, которые из них возникают.
В подтверждение своих взглядов Барнет ссылается на опыты, показавшие образование адаптивных ферментов плесенями и бактериями, «тренированными» к чужеродным субстратам.
Представление о существовании ферментов, обусловливающих образование специфических белков, как уже указывалось, не подтверждено никакими экспериментальными данными; однако в настоящее время мы не располагаем и такими экспериментальными данными, которые позволили бы окончательно его отвергнуть. Главное теоретическое возражение против этого утверждения состоит в том, что его вряд ли можно согласовать с современными физико-химическими представлениями о сущности катализа. Невозможно представить себе катализатор, который обусловливал бы синтез специфических пептидных цепей, соединяя аминокислоты в строго определенном порядке. Действие всех известных катализаторов и ферментов ограничено в каждый данный момент влиянием на одну определенную молекулярную группировку. После однократного выполнения своей функции фермент может повторно оказывать то же самое действие. Невозможно, однако, представить себе фермент, который катализировал бы, например, процесс присоединения тирозина к концевой аминной группе пептидной цепи, а затем способствовал бы присоединению аланина, глутаминовой кислоты, цистеина и других аминокислот. Все имеющиеся в нашем распоряжении данные о действии ферментов свидетельствуют о том, что фермент избирательно катализирует одну какую-либо реакцию и что характер его действия не меняется.
Выше, при обсуждении вопроса о механизме синтеза белков, мы говорили, с одной стороны, о роли белков как шаблонов для этого синтеза, с другой — о роли ферментов. Не нужно, однако, забывать, что ферменты также являются белками и что многие белки, которые считались лишенными ферментных функций, в действительности оказались ферментами, как только были найдены для них подходящие субстраты. Нортроп [150], доказавший, что ферменты образуются из проферментов, полагает, что и белки аналогичным путем могут образовываться из протеиногенов. Первой ступенью синтеза белков является, по Нортропу, образование из аминокислот типового белка — протеиногена, а следующей ступенью — аутокаталитическое образование отдельных белков из протеиногена [156].
В настоящее время, однако, нет никакой возможности критически обсудить эту точку зрения, поскольку нет никаких данных относительно механизма превращения профермента в фермент и протеиногена в белок.
Основная цель всех изложенных выше рассуждений о механизме синтеза белков состояла в том, чтобы показать, что физико-химическое объяснение этого процесса возможно согласовать с современными представлениями из области физической химии и биологии.
Это согласование достигнуто в данном случае путем устранения всего того, что противоречит установившимся физикохимическим концепциям или экспериментальным данным. Конечно, гораздо лучше было бы создать теорию синтеза белка, основываясь исключительно на фактах, полученных экспериментальным путем. В настоящее время, однако, это еще невозможно, вследствие чего приходится прибегать к различного рода гипотезам и предположениям.
Исследовательская работа немыслима без направляющих идей. Выдвижение новых гипотез допустимо поэтому там, где отсутствуют экспериментальные данные для построения достаточно обоснованных теорий.