ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
Клетка — это атом в биологии...Чтобы понять, что такое жизнь, нужно изучить клетку и определить ее строение с тем, чтобы познать закономерности жизнедеятельности любой клетки, найти те различия, которые отвечают за ее специфические функции.
Франсуа Жакоб, Логика жизни: история наследования, 1970
1. ОСНОВЫ БИОХИМИИ
Примерно пятнадцать-двадцать миллиардов лет назад в результате взрыва, сопровождавшегося извержением раскаленных субатомных частиц с очень высокой энергией, возникла Вселенная. Простейшие элементы (водород и гелий) образовались за считанные секунды. По мере расширения и остывания Вселенной материя под действием гравитации конденсировалась, и возникали звезды. Некоторые из них становились огромными и взрывались как сверхновые звезды, высвобождая энергию, необходимую для слияния простых атомных ядер и образования более сложных химических элементов. Так несколько миллиардов лет назад возникла Земля с теми химическими элементами, которые сейчас на ней существуют. Жизнь возникла около четырех миллиардов лет назад: появились простые микроорганизмы, способные добывать энергию из химических соединений, позже — из солнечного света и использовать ее для синтеза великого множества сложных биомолекул, используя простые элементы и соединения, находящиеся на поверхности Земли.
Биохимия изучает, каким образом замечательные свойства живых организмов возникают из тысяч различных неживых молекул. Выделенные в индивидуальном состоянии и исследованные, эти молекулы подчиняются всем физическим и химическим законам, описывающим поведение неживой материи; это справедливо для всех процессов, протекающих в живых организмах. Биохимические исследования позволяют понять, каким образом неживые молекулы в составе живых организмов взаимодействуют между собой, способствуя сохранению и непрерывному поддержанию жизни, причем и взаимодействия происходят строго в соответствии с физическими и химическими законами, управляющими неживой материей.
Тем не менее живые организмы обладают особыми признаками, отличающими их от других материальных объектов. Каковы же эти отличительные признаки?
Высокая сложность химической структуры и микроскопической организации. Тысячи различных молекул участвуют в составлении замысловатой внутренней структуры клетки (рис. 1-1, а). Среди них встречаются очень длинные полимеры (каждая макромолекула содержит характерный набор субъединиц и имеет уникальную трехмерную структуру), а также способность к высоко-специфическому, т. е. избирательному, связыванию с другими молекулами внутри клетки.
Система получения, преобразования и использования энергии окружающей среды (рис. 1-1, б) позволяет организму создавать и поддерживать свои внутренние структуры и выполнять механическую, химическую, осмотическую и электрическую работу. Это противодействует стремлению всей неживой материи к распаду и переходу в неупорядоченное состояние. В результате достигается равновесие с окружающей средой.
Рис. 1-1. Некоторые признаки живой материи, а) Электронная микрофотография окрашенного тонкого среза мышечной ткани позвоночного иллюстрирует сложность микроскопической организации живой ткани; б) степной сокол получает необходимые ему питательные вещества, поедая более мелких птиц; в) биологическое воспроизведение характеризуется изумительной точностью.

Способность к точному самовоспроизведению(рис. 1-1, в). Из единственной бактериальной клетки, помещенной в стерильную питательную среду, за 24 ч может возникнуть миллиард идентичных «дочерних» клеток. Каждая клетка содержит тысячи различных молекул, некоторые из них имеют чрезвычайно сложную структуру, но гем не менее каждая бактерия является точной копией оригинала, и се структура полностью определяется информацией, содержащейся в генетическом материале исходной клетки.
Механизмы восприятия изменений в окружающей среде непрерывно приводят организм в соответствие с условиями среды путем адаптации внутренней химической структуры или изменения положения в окружающей среде.
Специфические функции всех компонентов живого организма и регуляция их взаимоотношений. Это отличительная черта не только макроструктур, таких как стебель и лист или сердце и легкое, но также микроскопических внутриклеточных структур и индивидуальных химических соединений. Взаимодействие химических компонентов в живом организме динамично, изменения в одном соединении вызывают координирующие или компенсаторные изменения в других, причем вместе весь ансамбль приобретает черты, отличные от тех, что присущи его отдельным частям. Набор молекул выполняет некую программу, конечный результат которой состоит в воспроизведении этой программы и самого набора молекул, т. с. жизни.
Способность изменяться со временем путем последовательной эволюции. Организмы очень понемногу изменяют наследуемые стратегии выживания в соответствии с изменениями внешних условий. Результатом бесконечной эволюции является огромное многообразие форм жизни, внешне весьма различных (рис. 1-2), но в основе своей связанных общим происхождением. Эта общность всех живых организмов на молекулярном уровне выражается в сходстве последовательностей генов и структуры белков.
Рис. 1-2. Различные живые организмы имеют одинаковое химическое строение. Птицы, звери, растения, почвенные микроорганизмы и человек имеют одни и те же основные структурные единицы (клетки) и макромолекулы (ДНК, РНК, белки), построенные из мономерных единиц одинакового вида (нуклеотиды, аминокислоты). Живые организмы используют одни и те же метаболические пути для синтеза компонентов клетки, у них общий генетический код и они происходят от одних и тех же эволюционных предков. Приведен фрагмент картины Яна ван Кесселя мл. (1626-1679) «Сады Эдема».

Несмотря на эти общие свойства и основополагающее единство жизни, сложно сделать обобщения для всех живых организмов. Разнообразие жизни на Земле огромно. Диапазон условий существования организмов от горячих источников до арктической тундры и от кишечника животных до студенческих общежитии - соответствует широкому диапазону специфической биохимической адаптации, достигаемой в пределах общей химической структуры. Для большей ясности в данной книге мы иногда используем обобщения, которые, возможно, несовершенны, но полезны. Кроме того, мы часто обращаем внимание на исключения, которые можно подкрепить примерами.
Биохимия на молекулярном уровне описывает структуру, механизмы и химические процессы, свойственные всем организмам, и формулирует принципы организации, лежащие в основе любых форм жизни, которые можно назвать принципами молекулярной лотки жизни. Хотя биохимия вносит значительный вклад в фундаментальную и прикладную медицину, сельское хозяйство, систему питания и промышленность, ее основной задачей является изучение самой жизни.
В данной вступительной главе мы кратко остановимся па описании клеточных, химических, физических (термодинамических) и генетических основ биохимии и общего принципа эволюции - изменения свойств живых клеток на протяжении поколений. При прочтении книги возвращение к данной главе может оказаться полезным для восстановления в памяти изложенного здесь (фундаментального материала.
1.1. Принципы организации клетки
Единство и различие организмов становятся очевидными уже на клеточном уровне. Самые маленькие организмы состоят из одной клетки и имеют микроскопические размеры. Более крупные многоклеточные организмы содержат много разных типов клеток, отличающихся по размеру, форме п специфическим функциям. Несмотря на эти очевидные различия, все клетки как простейших, так и наиболее сложных организмов имеют общие фундаментальные свойства, которые можно исследовать па биохимическом уровне.
Клетки являются структурными и функциональными единицами всех живых организмов
Клетки всех видов имеют общие структурные особенности (рис. 1-3). Плазматическая мембрана ограничивает клетку, отделяя ее содержимое от окружающей среды. Мембрана состоит из молекул липидов и белков, образующих тонкий, плотный, пластичный, гидрофобный барьер вокруг клетки. Мембрана препятствует свободному проникновению в клетку неорганических ионов и большинства других заряженных или полярных соединений. Прохождение определенных ионов и молекул обеспечивают транспортные белки внутри плазматической мембраны; рецепторные белки передают в клетку сигналы; мембранные ферменты участвуют в некоторых метаболических реакциях. Поскольку отдельные липиды и белки в плазматической мембране не связаны между собой ковалентными связями, вся структура отличается замечательной гибкостью, позволяющей клетке изменять свою форму и размер. По мере роста клетки в плазматическую мембрану встраиваются вновь образующиеся молекулы липидов и белков. При делении каждой клетки образуются две новые, каждая из которых окружена собственной мембраной. Рост и деление (дробление) клетки происходят без нарушения целостности мембраны.
Рис. 1-3. Универсальные элементы строения живой клетки. Все клетки имеют ядро или нуклеоид плазматическую мембрану и цитоплазму. Цитозолем называют часть цитоплазмы, которая остается в супернатанте после центрифугирования клеточного экстракта при 150 000 д в течение 1 ч.
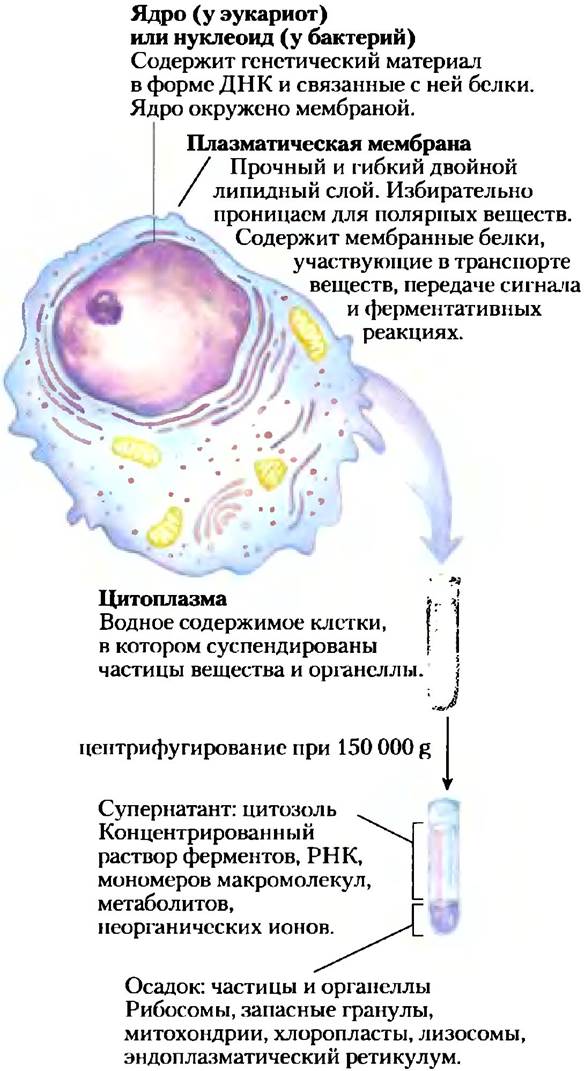
Внутреннее пространство, ограниченное плазматической мембраной, заполнено цитоплазмой (рис. 1-3), представляющей собой водную среду (цитозоль) с множеством частиц, выполняющих разнообразные функции. Цитозоль — это концентрированный раствор, содержащий ферменты и кодирующие их молекулы РНК, строительные блоки для этих макромолекул (аминокислоты и нуклеотиды), сотни небольших органических молекул (метаболитов — промежуточных продуктов биосинтеза и распада), коферментыдля многих ферментативных реакций, неорганические ионы и такие надмолекулярные структуры, как рибосомы, где происходит синтез белков, и протеасомы, где разрушаются те белки, в которых клетка больше не нуждается.
Все клетки, по крайней мере, в определенный период своей жизни имеют ядро или нуклеоцд, в котором хранится и реплицируется геном — полный набор генов, состоящих из ДНК. У бактерий и архей нуклеоид не отделяется от цитоплазмы мембраной. У большой группы эукариот (от греч. еu — истинный и karyon — ядро) ядро состоит из ядерного материала, заключенного в двухслойную мембрану — ядерную оболочку. Клетки, имеющие ядерную оболочку, называют эукариотическими. Все микроорганизмы, не имеющие ядерной оболочки, раньше называли прокариотами (от греч. pro — до, karyon — ядро), однако теперь среди них выделяют отдельно домен архей и домен бактерий (см. ниже).
Размеры клеток лимитированы диффузией кислорода
Большинство клеток имеют микроскопические размеры и невидимы невооруженным глазом. Диаметр клеток животных и растений — от 5 до 100 мкм, а длина большинства клеток одноклеточных микроорганизмов обычно не превышает 1-2 мкм. Что ограничивает размеры клетки? Нижний предел, по всей видимости, определяется минимальным количеством необходимых клетке биомолекул разных видов. Самые маленькие клетки бактерий — микоплазмы — имеют диаметр 300 нм и объем около 10-14 мл. Бактериальная рибосома в наибольшем измерении имеет размер 20 нм, следовательно, несколько рибосом занимают значительную часть объема клетки микоплазмы.
Верхний предел размера клетки, вероятно, определяется скоростью диффузии растворенных веществ в водной среде. Например, бактериальная клетка, использующая энергию реакций, протекающих с потреблением кислорода, получает молекулярный кислород, диффундирующий из окружающего пространства через плазматическую мембрану. Клетка настолько мала, а отношение площади ее поверхности к объему настолько велико, что диффундирующий кислород легко достигает любого участка цитоплазмы. Однако по мере увеличения размеров клетки снижается отношение площади ее поверхности к объему, и потребление кислорода в реакциях метаболизма увеличивается быстрее, чем количество кислорода, поступающего в клетку в результате диффузии. Таким образом, начиная с определенного размера клеток метаболизм с использованием О2 становится невозможным, что и определяет теоретический верхний предел размера клетки.
Существуют три царства живых организмов
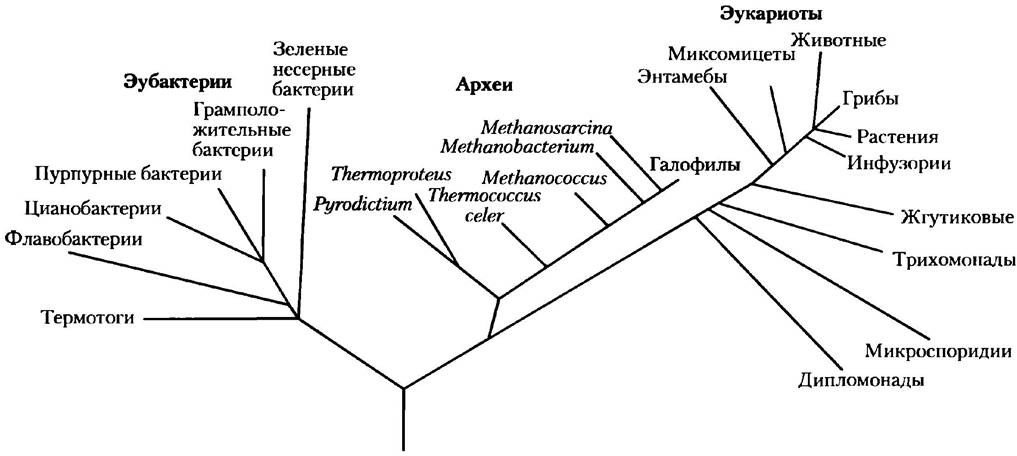
Каждый живой организм можно отнести к одной из трех больших групп (доменов) - трех ветвей эволюции, происходящих от общего предшественника (рис. 1-4). Две большие группы одноклеточных микроорганизмов различаются по своим биохимическим свойствам — это бактерии и археи. Бактерии населяют почву, поверхностные водоемы, а также ткани живых или разлагающихся организмов. Большинство наиболее хорошо изученных бактерий, в том числе Escherichia coli, относятся к эубактериям. Археи были выделены в отдельный домен Карлом Вёзе в 1980-х гг. Многие из этих организмов живут в экстремальных природных условиях, например, в соленых озерах, горячих источниках, болотах с очень высокой кислотностью воды и в глубинах океана. Существующие данные позволяют предположить, что археи и бактерии выделились в отдельные ветви на ранних этапах эволюции. Все эукариотические организмы составляют третий домен эукариот, который происходит из той же ветви эволюции, которая дала начало археям. Таким образом, археи являются более близкими родственниками эукариотам, чем бактериям.
Рис. 1-4. Филогения трех царств живых организмов. Филогенетическое родство часто иллюстрируют с помощью подобного «фамильного древа». Основой для построения этого древа является сходство нуклеотидных последовательностей рибосомной РНК внутри каждой группы; чем больше сходство последовательностей, тем ближе располагаются ветви, так что расстояние между двумя ветвями отражает степень расхождения двух последовательностей. Филогенетические деревья также могут быть основаны на сходстве последовательности аминокислот в отдельном белке. Например, на основе сравнения последовательностей белка GroEL (бактериальный белок, который участвует в сборке белков) построено древо, показанное на рис. 3-32. На рис. 3-33 представлено «консенсусное» древо, при построении которого для достижения более точных результатов эволюционного сродства групп организмов использованы несколько параметров сравнения, такие как вышеперечисленные.
Представителей доменов архей и бактерий разделяют на подгруппы в зависимости от условий их обитания. Аэробные организмы, населяющие места с достаточным содержанием кислорода, могут получать энергию от переноса электронов с топливных молекул на кислород. Микроорганизмы, адаптированные к анаэробным условиям, где кислород практически отсутствует, получают энергию путем переноса электронов на нитрат (с образованием N2), сульфат (с образованием Н2S) или СO2 (с образованием СН4). Многие организмы, эволюционировавшие в анаэробных условиях, являются облигатными (строгими) анаэробами: в присутствии кислорода они погибают. Другие —факультативные анаэробы, они могут жить и без кислорода, и в его присутствии.
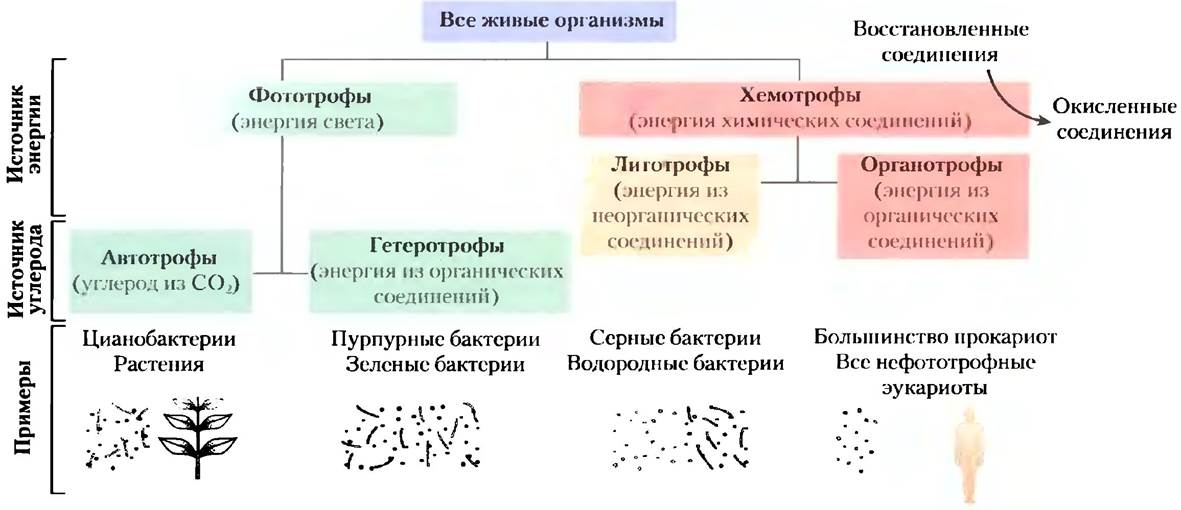
Организмы можно классифицировать в соответствии с тем способом, с помощью которого они получают энергию и углерод, необходимые для синтеза клеточного вещества (рис. 1-5). В зависимости от источника энергии выделяют две большие группы организмов: фототрофы (от греч. trophe — питание) используют солнечный свет, а хемотрофы добывают энергию путем окисления топливных молекул. Группа хемотрофных организмов, называемых литотрофами, способна окислять неорганические вещества: НS- до элементарной серы S0, S0 до SO42-, NO2- до NO3, а Fe2+ до Fe3+. Органотрофы окисляют различные органические вещества, находящиеся в окружающей их среде. Фототрофы и хемотрофы также можно разделить на группы в соответствии с тем, могут ли они использовать в качестве источника углерода СO2 (автотрофы) или должны получать углерод из органических питательных веществ (гетеротрофы).
Рис. 1-5. Организмы можно классифицировать в соответствии с теми источниками энергии (солнечный свет либо окисление химических соединений) и углерода, которые они используют для синтеза клеточных веществ.
Бактерия Escherichia coli — наиболее изученная бактерия
Строение клетки бактерий имеет общие структурные черты, а также некоторые групповые особенности (рис. 1-6). В нормальных условиях Е. coli является безопасным обитателем кишечника человека. Длина ее клетки около 2 мкм, диаметр чуть менее 1 мкм. Клетка защищена внешней мембраной, а внутренняя плазматическая мембрана ограничивает пространство, в котором заключены цитоплазма и нуклеоид. Пространство между внешней и внутренней мембранами занято тонким, но прочным слоем полимеров, называемых пептидогликанами, которые придают клетке жесткость и определенную форму. Плазматическая мембрана вместе с внешними по отношению к ней слоями образует клеточную оболочку. Прочность клеткам архей придают полимеры другого типа — псевдопептидогликаны. Плазматическая мембрана бактерий состоит из тонкого двойного слоя липидов с белковыми включениями. Мембраны архей имеют аналогичное строение, однако их липиды поразительным образом отличаются от липидов бактерий (см. рис. 10-12).
Рис. 1-6. Общие структурные элементы бактериальных клеток. В связи с различным строением клеточной оболочки некоторые эубактерии (грамположительные бактерии) удерживают краситель при окрашивании по Граму, а некоторые — нет (грамотрицательные бактерии). Е. coli относится к грамотрицательным бактериям. Цианобактерии представляют собой эубактерии, но отличаются особой структурой внутренней мембраны, в которой локализованы фотосинтетические пигменты. Клеточные оболочки архей и грамположительных эубактерий под электронным микроскопом выглядят сходным образом, однако строение их мембранных липидов и полисахаридов заметно различается (см. рис. 10-12).
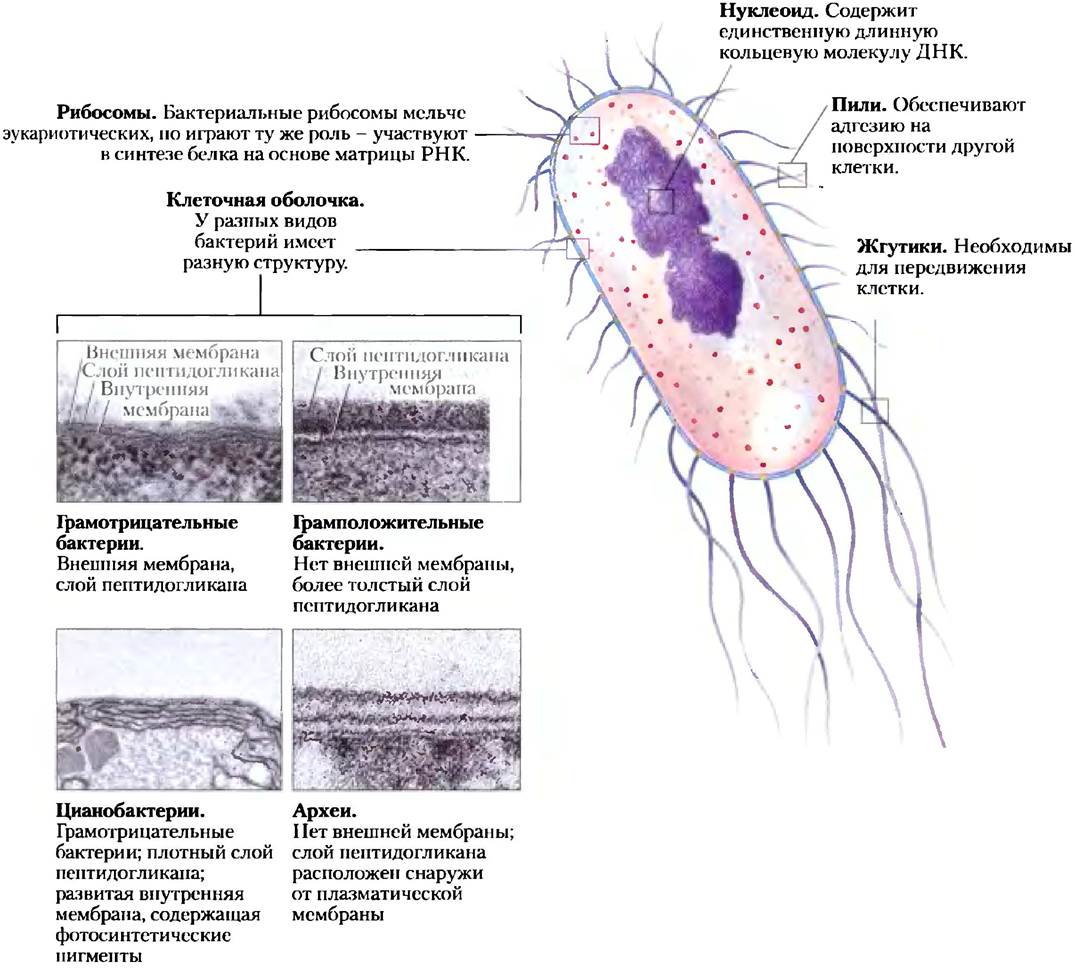
В цитоплазме Е. coli содержится около 15 000 рибосом, от десяти до тысячи копий каждого из примерно 1000 различных ферментов, а также около 1000 органических веществ с молекулярной массой менее 1000 (метаболиты и кофакторы). В нуклеоиде расположена единственная кольцевая молекула ДНК, а в цитоплазме, как и у многих других бактерий, встречается одна или несколько более мелких кольцевых молекул ДНК, называемых плазмидами. В природных условиях некоторые плазмиды обеспечивают устойчивость бактерий к различным находящимся в окружающей среде токсинам и антибиотикам. В лабораторных условиях эти молекулы ДНК необычайно удобны для осуществления экспериментальных манипуляций и очень широко используются молекулярными генетиками.
Большинство бактерий (включая Е. coli) существуют в виде индивидуальных клеток, но некоторые виды бактерий (например, миксобактерии) демонстрируют простейшее «социальное поведение», образуя многоклеточные агрегаты.
Эукариотические клетки содержат разнообразные мембраносвязанные органеллы, которые можно выделить и исследовать
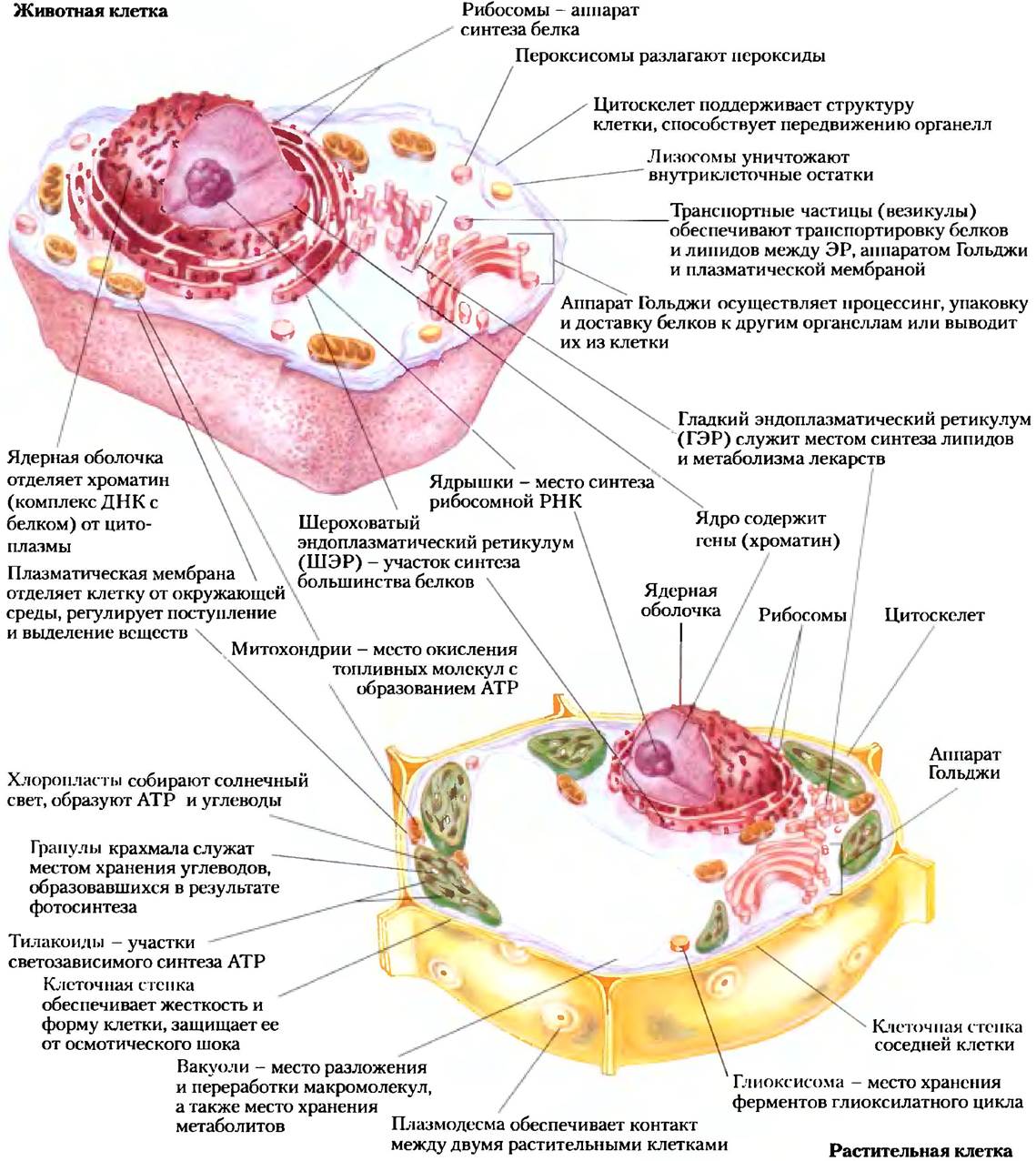
Типичные эукариотические клетки (рис. 1-7) во много раз превышают по размеру бактериальные и обычно имеют диаметр от 5 до 100 мкм, а их объем в тысячи или миллионы раз больше объема бактериальных клеток. Отличительной особенностью эукариот является наличие ядра и многочисленных связанных с мембраной органелл со специфическими функциями: митохондрий, эндоплазматического ретикулума, комплекса Гольджи, пероксисом и лизосом. В растительных клетках, кроме того, содержатся вакуоли и хлоронласты (рис. 1-7). В цитоплазме многих клеток присутствуют [ранулы или капельки, содержащие запасные питательные вещества, такие как крахмал и жиры.
Рис. 1-7. Структура эукариотической клетки. Схематически изображены клетки двух основных типов: вверху — животная клетка, внизу — растительная клетка. Клетки растений обычно имеют диаметр от 10 до 100 мкм, а животные клетки — от 5 до 30 мкм. Структуры, выделенные красным цветом, присутствуют только в животных или только в растительных клетках. В клетках эукариотических микроорганизмов (например, протистов и грибов) имеются такие же структуры, как в клетках растений и животных, а кроме того, во многих из них также есть специализированные органеллы, которые здесь не изображены.
Значительный вклад в развитие биохимии внесли работы Альбера Клода, Кристиана де Дюва и Джорджа Паладе, посвященные методам разделения клеточных органелл — важного этапа в выделении и изучении функций биомолекул и более крупных клеточных структур. При обычной процедуре фракционирования (рис. 1-8) клетки или ткани, находящиеся в растворе, гомогенизируют в мягких условиях. При этом происходит разрыв плазматической мембраны, но большинство клеточных органелл сохраняют свою целостность. Затем гомогенат центрифугируют. Ядра, митохондрии и лизосомы различаются по размеру и поэтому имеют различную скорость седиментации. Они также различаются по удельной плотности и при центрифугировании в градиенте плотности оказываются в разных фракциях.
Рис. 1-8. Разделение субклеточных структур ткани. Для получения внутриклеточных органелл ткани, например, печены сначала подвергают механической гомогенизации для разрушения клеток и диспергирования их содержимого в буфере. Раствор сахарозы обеспечивает практически такое же осмотическое давление, какое существует внутри клеточных органелл, что препятствует проникновению в органеллы воды, их разбуханию и разрушению, а) Крупные и мелкие частицы в суспензии можно разделить центрифугированием с разными скоростями. б) Частицы различной плотности можно разделить изопикническим центрифугированием. В данном методе центрифужные пробирки заполняют раствором, плотность которого возрастает сверху вниз. Для получения градиента плотности готовят, например, растворы с разной концентрацией сахарозы. Затем смесь органелл помещают в верхнюю часть пробирки и центрифугируют с высокой скоростью. При этом органеллы осаждаются в градиенте до того уровня, плотность которого точно соответствует их собственной. После центрифугирования каждый слой можно отобрать из пробирки отдельно.
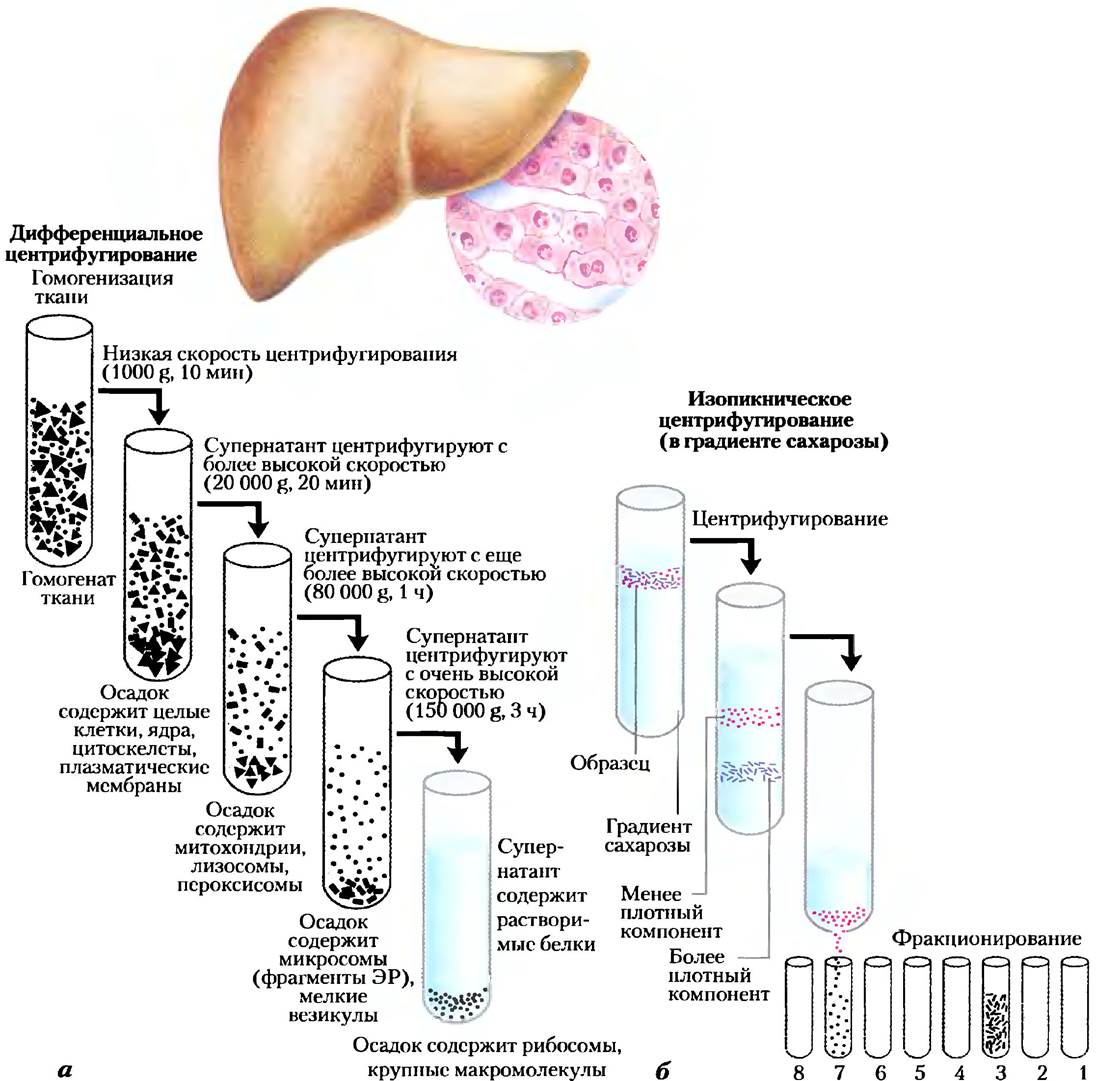
Дифференциальное центрифугирование приводит к грубому фракционированию содержимого цитоплазмы. Далее фракции можно разделять путем изоникнического* центрифугирования. При этом органеллы с различной плавучей плотностью (результат различного соотношения в них белков и липидов) разделяются центрифугированием в слое растворителя с градиентом плотности. Осторожно изолируя содержимое каждого участка градиента и рассматривая его в микроскоп, биохимик может установить положение фракции каждой органеллы в центрифужной пробирке и выделить очищенную фракцию для дальнейшего изучения. С использованием этого метода удалось установить, например, что лизосомы содержат гидролитические ферменты, митохондрии — окислительные ферменты, а хлоропласты — фотосинтетические пигменты. Выделение органеллы, содержащей определенный фермент, часто является первой стадией очистки данного фермента.
* Изоникнический — имеющий ту же плотность.
Цитоплазма имеет цитоскелет и обладает динамическими свойствами
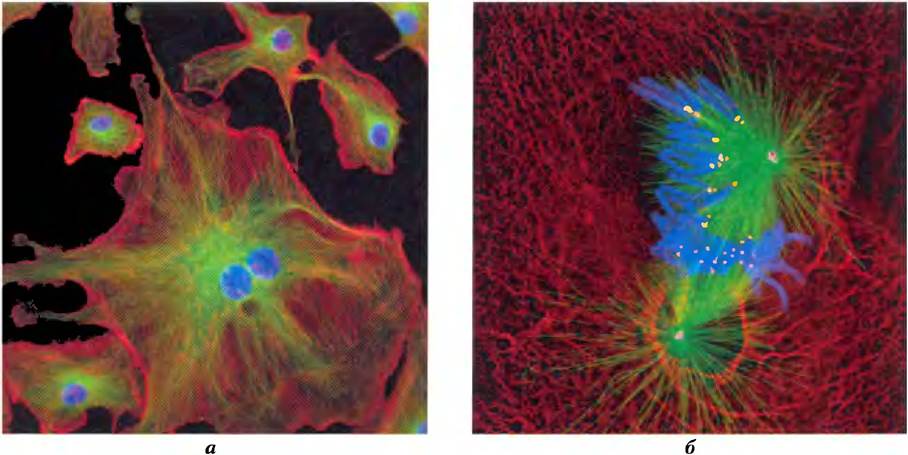
С помощью флуоресцентной микроскопии можно различить несколько типов белковых филаментов, пересекающих крест-накрест эукариотическую клетку и образующих трехмерную сеть, называемую цитоскелетом. Существуют три основных типа цитоплазматических филаментов — актиновые филаменты, микротрубочки и промежуточные филаменты (рис. 1-9); они различаются по толщине (от 6 до 22 нм), строению и специфическим функциям. Все филаменты служат для организации и структурирования цитоплазмы и придают форму клетке. Актиновые филаменты и микротрубочки также способствуют передвижению органелл и целых клеток.
Все филаменты цитоскелета, независимо от типа, построены из простых белковых молекул, которые, связываясь друг с другом нековалентно, образуют тяжи определенной толщины. Эти филаменты не являются постоянными структурами: они непрерывно диссоциируют на составляющие их единицы и собираются вновь. Их положение в клетке также не фиксировано строго, а может значительно изменяться при митозе, цитокинезе, амебоидном движении или при изменениях формы клетки. Разборка, сборка и локализация всех типов филаментов регулируются другими белками, которые необходимы для связывания филаментов или перемещения вдоль них цитоплазматических органелл.
Рис. 1-9. Три типа волокон цитоскелета: актиновые волокна, микротрубочки и промежуточные волокна. Клеточные структуры можно пометить с помощью антител (узнающих определенные белки) с флуоресцентной меткой, присоединенной ковалентной связью. Если обработанные таким образом клетки разглядывать с помощью флуоресцентного микроскопа, то соответствующие структуры становятся видимыми, а) Клетки эндотелия легочной артерии быка. В красный цвет окрашены пучки актиновых волокон, называемые стрессовыми волокнами; в зеленый цвет окрашены выходящие из центра клетки микротрубочки; в синий цвет — хромосомы внутри ядра. 6) Клетки легких тритона в митозе. Микротрубочки (зеленый цвет), прикрепленные к кинетохорам (желтый цвет) на конденсированных хромосомах (синий цвет), растягивают хромосомы к противоположным полюсам клетки (центросомы; малиновый цвет). Промежуточные волокна, построенные из кератина (красный цвет), служат для поддержания структуры клетки.
Описанная выше структура соответствует ситуации, наблюдающейся в эукариотической клетке с сетчатой структурой нитей и сложной организаций мембрапосвязанныхкомпартментов (рис. 1-7). Филаменты разбираются и вновь собираются в каком-то другом участке клетки. Окруженные мембраной частицы отпочковываются от одной органеллы и сливаются с другой. Органеллы переметаются в цитоплазме вдоль белковых филаментов, энергию для этих передвижений обеспечивают моторные белки. Внутренние мембраны разделять отдельные метаболические процессы и служат поверхностями, на которых протекают некоторые ферментативные реакции. Транспортные механизмы эндо- и экзоцитоза, способствующие проникновению веществ соответственно в клетку и из нее, вязаны со слиянием и раскрытием мембран. Эти процессы обеспечивают обмен между цитоплазмой и окружающей средой и позволяют осуществлять секрецию произведенных в клетке продуктов и захват внеклеточных веществ.
Подобная организация цитоплазмы сложна, но далеко не хаотична. Движение и локализация органелл и элементов цитоскелета находятся под строгим контролем. На определенных этапах жизни эукариотической клетки происходят серьезные, точно спланированные перестройки, такие как митоз. Взаимодействия между цитоскелетом и органеллами являются нековалентными и обратимыми; они регулируются в ответ на различные внутри- и внеклеточные сигналы.
В клетках существуют надмолекулярные структуры
Макромолекулы и их мономерные звенья сильно различаются по размерам (рис. 1-10). Молекула аланина имеет длину менее 0,5 нм. Переносящая кислород молекула гемоглобина в эритроцитах состоит приблизительно из 600 аминокислотных звеньев, организованных в четыре длинные цепи, сложенные в глобулы; в результате нативный гемоглобин достигает в диаметре 5,5 нм. Белки, в свою очередь, гораздо мельче рибосом (диаметр последних около 20 нм), а те значительно уступают по размерам таким органеллам, как митохондрии (диаметр митохондрий около 1000 нм). Существует колоссальная разница между простыми биомолекулами и теми клеточными структурами, которые можно наблюдать в световой микроскоп. Иерархия в структурной организации клетки изображена на рис. 1-11.
Рис. 1-10. Органические молекулы, из которых построена основная часть клеточного вещества: биохимический алфавит, а) Шесть из двадцати аминокислот, из которых состоят все белки (боковая цепь выделена розовым цветом); б) пять азотистых оснований, два пятиуглеродных сахара и фосфорная кислота, из которых построены все нуклеиновые кислоты; в) пять компонентов мембранных липидов; г) D-глюкоза — мономерное звено большинства углеводов. Заметьте, что фосфорная кислота входит в состав как нуклеиновых кислот, так и мембранных липидов.
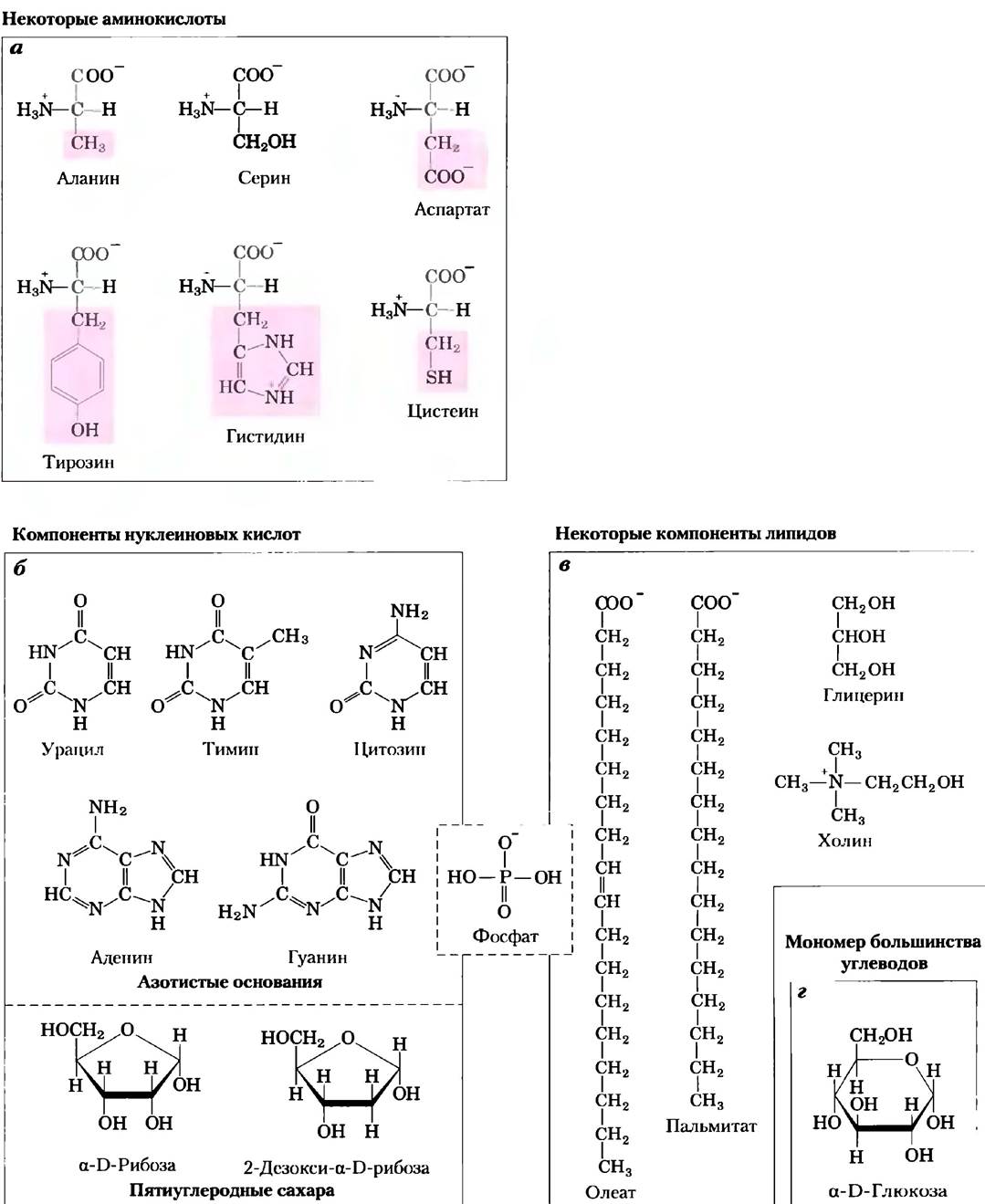
Рис. 1-11. Структурная иерархия в молекулярной организации клетки. В данной растительной клетке ядро содержит несколько типов надмолекулярных комплексов, включая хромосомы. Хромосомы состоят из макромолекул ДНК и множества различных белков. Каждый тип макромолекул построен из простых мономерных звеньев, например, ДНК состоит из нуклеотидов (дезоксирибонуклеотидов).
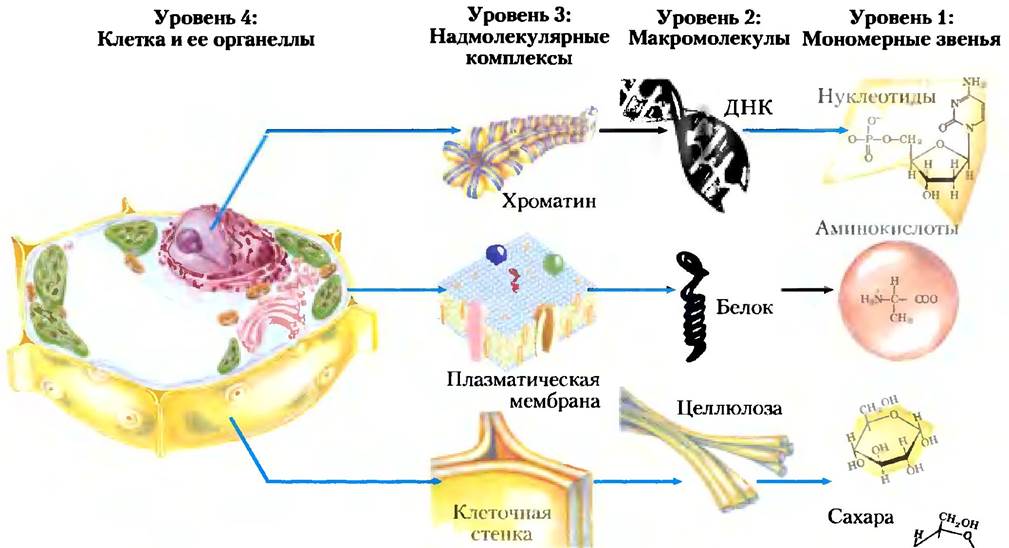
Мономерные звенья белков, нуклеиновых кислот и полисахаридов связаны ковалентными связями. В надмолекулярных комплексах молекулы удерживаются вместе гораздо более слабыми нековалентными взаимодействиями, среди которых выделяют водородные связи (между полярными группами), ионные (между заряженными группами), гидрофобные (между неполярными группами в водных средах) и ван-дер-ваальсовы взаимодействия, причем все они обладают гораздо меньшей энергией, чем ковалентная связь.
Природа этих нековалентных взаимодействий обсуждается в гл. 2. Макромолекулы в надмолекулярных структурах удерживаются большим числом слабых взаимодействий, обеспечивающих уникальную структуру этих комплексов.
В исследованиях in vitro можно не заметить важных взаимодействий между молекулами
Одним из подходов к исследованию биологических процессов является изучение очищенных молекул in vitro («в пробирке») при отсутствии других «лишних» молекул, находящихся в интактной (т. е. in vivo — «в живом организме») клетке. Такой подход весьма полезен, однако нс следует забывать, что содержимое клетки очень сильно отличается от содержимого пробирки. «Лишние» компоненты, удаленные при очистке, могут играть важнейшую роль для биологического действия или для регуляции выделенной молекулы. Например, изучение чистых ферментов in vitro обычно проводят при очень низких концентрациях ферментов в тщательно перемешиваемых водных растворах. В клетках ферменты растворены или суспендированы в желеобразном цитозоле вместе с тысячами других белков, некоторые из которых связывают данный фермент и влияют па его активность. Многие ферменты входят в полиферментные комплексы, в которых реагирующие вещества «передаются» от одних ферментов к другим, вовсе не выходя в окружающую среду. В желеобразном цитозоле диффузия затруднена, поэтому его состав в разных участках клетки может быть различным. Короче говоря, исследуемая молекула может вести себя в пробирке совсем не так, как в клетке. Основной задачей биохимии является изучение влияния клеточной организации и макромолекулярных комплексов на функционирование отдельных ферментов и других биологических молекул для понимания законов их функционирования как in vivo, так и invitro.
Краткое содержание раздела 1.1. Принципы организации клетки
■ Все клетки ограничены плазматической мембраной, имеют цитозоль, в котором растворены метаболиты, коферменты, неорганические ионы и ферменты, а также содержат набор генов, заключенных в нуклеоид (бактерии и археи) или ядро (эукариоты).
■ Фототрофы для своей жизнедеятельности используют солнечный свет, хемотрофы окисляют топливные молекулы, передавая электроны на акцепторы электронов: неорганические вещества, органические вещества или молекулярный кислород.
■ Бактериальные клетки и клетки архей имеют цитозоль, нуклеоид и плазмиды. Эукариотические клетки содержат ядро и разделены на множество компартментов, где в различных органеллах протекают специфические процессы; органеллы можно выделить и изучать отдельно.
■ Белки цитоскелета собраны в длинные нити (филаменты), придающие клетке прочность, определяющие ее форму и служащие в качестве направляющих путей для движения клеточных органелл.
■ Надмолекулярные комплексы удерживаются за счет нековалентных взаимодействий и имеют иерархическую структуру; некоторые комплексы видны в световой микроскоп. При выделении отдельных молекул из этих комплексов для исследований in vitro могут быть упущены некоторые важные закономерности жизнедеятельности клетки.