Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Биосинтез углеводов в животных тканях
В рассмотрении клеточного метаболизма мы достигли теперь как бы поворотного пункта. До сих пор мы знакомились с тем, как главные типы питательных веществ-углеводы, жирные кислоты и аминокислоты,- расщепляясь, включаются по сходящимся катаболическим путям в цикл лимонной кислоты, чтобы передать свои богатые энергией электроны в дыхательную цепь. Перемещаясь по дыхательной цепи к кислороду, эти электроны поставляют энергию для синтеза АТР. Теперь нам предстоит рассмотреть анаболические пути. На этих путях химическая энергия в форме АТР и NADPH используется для синтеза клеточных компонентов из простых предшественников. Катаболизм и анаболизм протекают одновременно; при этом поддерживается динамическое стационарное состояние, так что расщепление клеточных компонентов, обеспечивающее клетки энергией, уравновешивается биосинтетическими процессами, которые создают и поддерживают в живых клетках присущую им упорядоченность.
Здесь уместно вспомнить (гл. 10) и лишний раз подчеркнуть некоторые организационные принципы биосинтеза.
1. Пути биосинтеза и пути расщепления тех или иных биомолекул, как правило, не идентичны. Эти пути могут включать какую-нибудь общую обратимую реакцию или даже несколько таких реакций, но у них всегда имеется хотя бы одна ферментативная стадия, по которой они различаются. Если бы катаболические и анаболические реакции катализировались одним и тем же набором ферментов, действующих обратимо, то никакая биологическая структура независимо от ее сложности попросту не могла бы существовать, потому что число клеточных макромолекул менялось бы в ответ на любые колебания концентраций молекул-предшественников.
2. Биосинтетические пути и соответствующие им катаболические пути контролируются разными регуляторными ферментами. Обычно регуляция соответствующих биосинтетических и катаболических путей осуществляется координированным образом, реципрокно, так что стимулирование биосинтетического пути сопровождается подавлением катаболического пути, и наоборот. Более того, биосинтетические пути регулируются обычно на одном из первых этапов. Это избавляет клетку от непроизводительных трат: она не расходует предшественники на синтез тех промежуточных продуктов, которые ей не понадобятся. Мы вновь убеждаемся на этом примере, что принцип экономии лежит в основе молекулярной логики живых клеток.
3. Требующие затраты энергии биосинтетические процессы обязательно сопряжены с поставляющим энергию расщеплением АТР, вследствие чего весь процесс в целом является практически необратимым, точно так же как в целом необратим катаболизм. Таким образом, общее количество энергии АТР (или NADPH), используемое на данном биосинтетическом пути, всегда превосходит то минимальное количество свободной энергии, которое требуется для превращения предшественника в биосинтетический продукт.
Рассмотрение биосинтетических процессов мы начнем с центрального биосинтетического пути, который в животных тканях приводит к образованию различных углеводов из неуглеводных предшественников. У всех высших животных биосинтез D-глюкозы - абсолютно необходимый процесс, потому что D-глюкоза крови служит единственным или главным источником топлива для нервной системы (в том числе и для мозга), а также для почек, семенников, эритроцитов и для всех тканей эмбриона. У человека один только мозг потребляет более 120 г глюкозы в сутки. В организме животных D-глюкоза непрерывно синтезируется в строго регулируемых реакциях из более простых предшественников, таких, как пируват и некоторые аминокислоты, а затем поступает в кровь. Из неуглеводных предшественников образуются также и другие важные углеводы (рис. 20-1). Особенно большое значение имеет биосинтез гликогена, протекающий в печени и мышцах. Гликоген печени служит резервуаром глюкозы: из него образуется глюкоза, которая поступает в кровь. Мышечный же гликоген, распадаясь в процессе гликолиза, служит источником энергии АТР для мышечного сокращения. У животных образование D-глюкозы из неуглеводных предшественников называют глюконеогенезом (образование «нового» сахара). Важными предшественниками D-глюкозы являются у них лактат, пируват, глицерол, большинство аминокислот и промежуточные продукты цикла лимонной кислоты (рис. 20-1). Глюконеогенез протекает у животных главным образом в печени и значительно менее интенсивно - в корковом веществе почек.
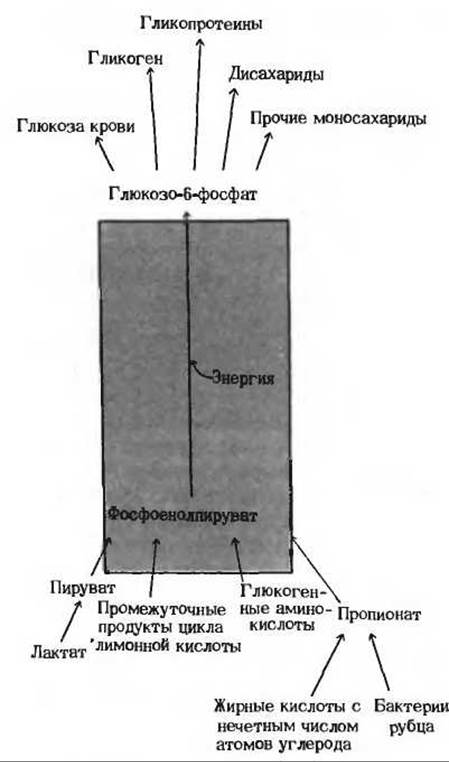
Рис. 20-1. Путь, ведущий от фосфоенолпирувата к глюкозо-6-фосфату, является общим для превращения многих предшественников в различные углеводы в животных тканях.
Мы знаем, что в растительном мире огромные количества глюкозы, а также других углеводов образуются путем восстановления двуокиси углерода в процессе фотосинтеза (гл. 23). В отличие от растений у животных не происходит реального (net) превращения СО2 в новые молекулы глюкозы.