Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Метаболизм. Общий обзор
Соответствующие катаболические и анаболические пути различаются, и эти различия имеют важное значение
Катаболический путь и соответствующий ему, но противоположный по направлению анаболический путь между данным предшественником и данным продуктом обычно не совпадают. Могут различаться и промежуточные продукты, и отдельные стадии этих путей. Например, протекающее в печени расщепление глюкозы до пирувата представляет собой процесс, состоящий из 11 последовательных стадий, катализируемых специфичными ферментами. Казалось бы, синтез глюкозы из пирувата должен быть простым обращением всех этих ферментативных стадий ее распада; такой путь представляется на первый взгляд и самым естественным, и наиболее экономичным. Однако в действительности биосинтез глюкозы в печени протекает иначе. Он включает лишь 9 из 11 ферментативных стадий, участвующих в ее распаде, а две недостающие стадии заменены в нем совсем другим набором ферментативных реакций, свойственным одному только биосинтетическому пути. Точно так же неидентичны и соответствующие пути катаболизма и анаболизма, связывающие, например, белки с аминокислотами или жирные кислоты с ацетил-СоА.
Можно было бы счесть ненужным расточительством наличие двух отдельных метаболических путей между двумя данными пунктами. Есть, однако, важные причины для того, чтобы катаболические и анаболические пути не совпадали. Первая из них заключается в том, что путь, по которому идет расщепление той или иной биомолекулы, может быть непригодным для ее биосинтеза по энергетическим соображениям. Расщепление какой- нибудь сложной органической молекулы можно сравнить со спуском с горы, а ее биосинтез - с подъемом в гору; в первом случае свободная энергия выделяется, а во втором ее требуется затратить, чтобы осилить подъем. Попробуем пояснить это с помощью простой аналогии. Если столкнуть с вершины горы валун, то он покатится вниз, теряя при этом энергию. На некоторых, особо крутых участках пути, при отвесном падении, теряются сразу большие количества энергии. Втащить валун трактором на вершину по тому же пути, по которому он скатился вниз, скорее всего не удастся. Трактор сможет, вероятно, подняться вверх по более пологой дороге, минуя крутые склоны (рис. 13-8). На этот обходный путь потребуется дополнительная энергия. Биосинтетический путь тоже требует дополнительных затрат энергии на преодоление крутых участков энергетической «горки».
Вторая причина, по которой соответствующие катаболические и анаболические пути неидентичны, состоит в том, что эти последовательности реакции должны регулироваться раздельно. Если бы для расщепления и для биосинтеза использовался один и тот же путь, т. е. имело бы место простое обращение последовательности реакций, то, например, торможение катаболического пути вследствие ингибирования одного из его ферментов неизбежно влекло бы за собой также и замедление соответствующего биосинтетического пути. Для того чтобы синтез и распад какого-либо соединения могли регулироваться независимо друг от друга, эти метаболические пути должны быть совершенно различными, а если у них все же имеются какие-то общие ферментативные стадии, то регулировать скорость процесса должны те ферменты, которые в противоположной последовательности реакций не участвуют (рис. 13-9).
Иногда противоположно направленные катаболические и анаболические пути различаются по своей локализации. Так, например, окисление жирных кислот до стадии ацетил-СоА в печени катализируется набором ферментов, локализованных по преимуществу в митохондриях, где условия благоприятствуют окислению; синтез же жирных кислот из ацетил-СоА, требующий водородных атомов, т.е. восстановительной способности, осуществляется с помощью другого набора ферментов, локализованных в цитозоле, где условия благоприятствуют восстановительным реакциям (рис. 13-10).
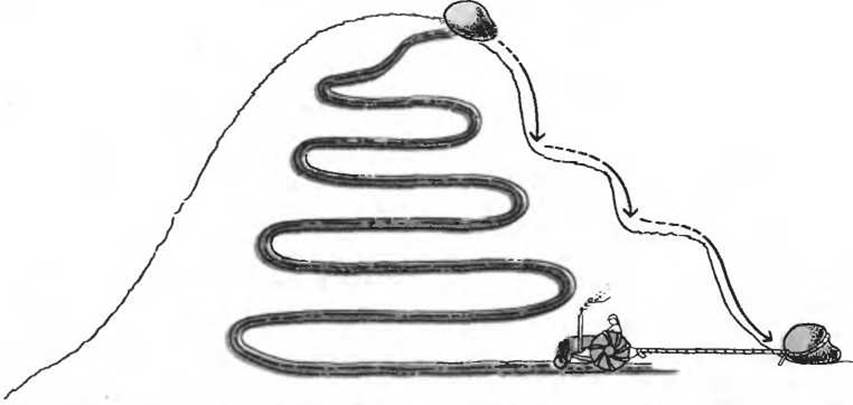
Рис. 13-8. Аналогия, поясняющая энергетические аспекты катаболизма и анаболизма на примере скатывающегося с горы валуна. Катаболизм можно сравнить со спуском с горы, так как он сопровождается потерей свободной энергии. Особенно много энергии теряется на крутых, почти отвесных участках пути (обозначены стрелками). Анаболизм напоминает подъем в гору; он требует затраты свободной энергии, которая может поступать лишь небольшими, строго определенными порциями. Трактор, например, смог бы втащить валун обратно на вершину горы только при условии, что он пройдет другим, более пологим путем, минуя крутые участки, на преодоление которых потребовалось бы слишком много энергии.
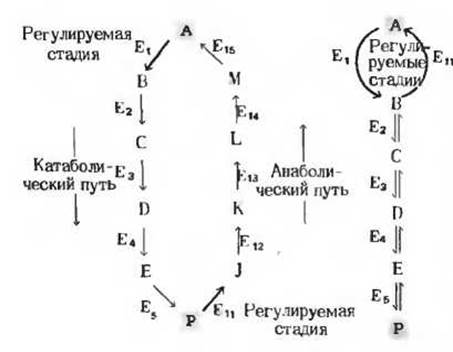
Рис. 13-9. Параллельные катаболические и анаболические пути должны быть различными хотя бы на одной из ферментативных стадий, для того чтобы они могли регулироваться независимо. Показаны два варианта независимой регуляции катаболического и анаболического путей между А и Р. В первом варианте эти пути совершенно различны, т.е. катализируются разными наборами ферментов. Во втором анаболический и катаболический пути различаются лишь по одному ферменту. Регулируемые стадии в обоих вариантах обозначены красными стрелками.
Однако хотя соответствующие катаболические и анаболические пути неидентичны, их связывает общая стадия (стадия III на рис. 13-6), которая включает в себя цикл лимонной кислоты и некоторые вспомогательные ферментативные реакции. Эту общую стадию называют иногда амфиболической стадией метаболизма (от греч. «аmfi»-оба), поскольку она выполняет двойную функцию. В катаболизме на этой стадии завершается распад сравнительно небольших молекул, образовавшихся на стадии II, а в анаболизме ее роль заключается в поставке небольших молекул-предшественников для биосинтеза аминокислот, жирных кислот и углеводов (об этом мы еще будем говорить далее).
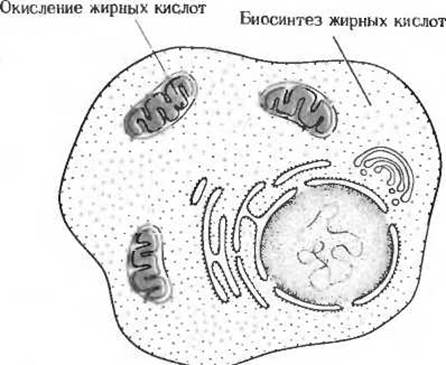
Рис. 13-10. Пространственное разделение противоположно направленных метаболических путей. Окисление жирных кислот происходит в основном в митохондриях, между тем как их синтез, для которого требуется восстановительная способность, протекает в цитозоле.
Почти все метаболические реакции в конечном счете связаны между собой, поскольку продукт одной ферментативной реакции служит субстратом для другой, которая в данном процессе играет роль следующей стадии. Таким образом, мы можем представить себе метаболизм в виде чрезвычайно сложной сети ферментативных реакций. Если поток питательных веществ в какой-нибудь одной части этой сети уменьшится или нарушится, то в ответ могут произойти изменения в другой части сети, для того чтобы это первое изменение было как-то уравновешено или скомпенсировано. На любом из центральных метаболических путей - катаболическом или анаболическом - скорость может регулироваться в соответствии с сиюминутными потребностями клетки. Более того, и катаболические, и анаболические реакции отрегулированы, по-видимому, таким образом, чтобы они протекали наиболее экономно, т.е. с наименьшей возможной затратой энергии и веществ. Например, окисление питательных веществ в клетке совершается со скоростью, как раз достаточной для того, чтобы удовлетворить ее энергетические потребности в данный момент.