Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
ДНК: структура хромосом и генов
Нуклеотидные последовательности некоторых ДНК уже расшифрованы
В 1977 г. была определена полная нуклеотидная последовательность ДНК бактериофага фХ174 (с. 850). Это выдающееся достижение ознаменовало собой начало новой эры в биохимии генов и хромосом. С тех пор была расшифрована нуклеотидная последовательность целого ряда генов, и в настоящее время, по крайней мере в принципе, можно определить последовательность оснований, по- видимому, любой ДНК.
До 1977 г. была расшифрована нуклеотидная последовательность многих тРНК и нескольких небольших мРНК. Роберт Холли и его коллеги первыми определили нуклеотидную последовательность нуклеиновой кислоты-дрожжевой аланиновой тРНК. Это исследование, завершенное в 1965 г., потребовало нескольких лет работы. Хотя молекулы тРНК состоят менее чем из 100 нуклеотидных остатков, они содержат много необычных модифицированных оснований, которые необходимо было идентифицировать. При секвенировании (т.е. определении нуклеотидной последовательности) ДНК возникают также и другие трудности. Мы уже упоминали, что в среднем ген Е. coli состоит из 1200 нуклеотидных пар, а целая молекула ДНК бактериофага фХ174 - более чем из 5000 пар. Раньше не было методов, позволяющих избирательно расщеплять ДНК по определенному нуклеотиду - скажем, по всем остаткам А. Даже если бы такой метод существовал, при расщеплении образовался бы очень большой набор фрагментов меньшего размера, которые чрезвычайно трудно было бы разделить. Более того, если бы даже удалось разделить и секвенировать эти фрагменты, восстановить из них полную последовательность оказалось бы практически невозможным.
Решающий успех был достигнут благодаря трем основным достижениям. Первым из них явилось открытие рестриктирующих эндонуклеаз, которые расщепляют молекулы ДНК только в сравнительно небольшом числе специфических точек. Использование двух или большего числа рестриктирующих эндонуклеаз (табл. 27-7) позволило расщеплять молекулы ДНК на отдельные фрагменты разными способами с образованием перекрывающихся последовательностей - точно так же, как применение двух различных протеолитических ферментов (например, трипсина и химотрипсина) открыло в свое время возможность расщеплять полипептидные цепи на разные наборы фрагментов и устанавливать аминокислотные последовательности в перекрывающихся участках (разд. 6.7, е). Например, ДНК обезьяньего вируса 40 (SV40) (рис. 27-30), способного превращать некоторые клетки в злокачественные, была расщеплена рестриктирующими эндонуклеазами в ряде специфических точек с образованием фрагментов, удобных для определения локализации отдельных генов. На рис. 27-31 схематически показаны сайты расщепления ДНК SV40 с помощью трех рестриктирующих эндонуклеаз.
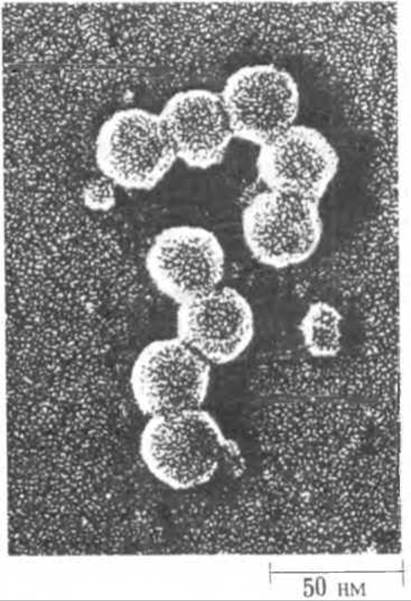
Рис. 27-30. Обезьяний вирус 40 (SV40) вызывает рак у хомячков и других мелких животных. Он представляет собой один из самых мелких канцерогенных вирусов, Белковая оболочка SV40 имеет форму икосаэдра, т.е. двадцатигранника.
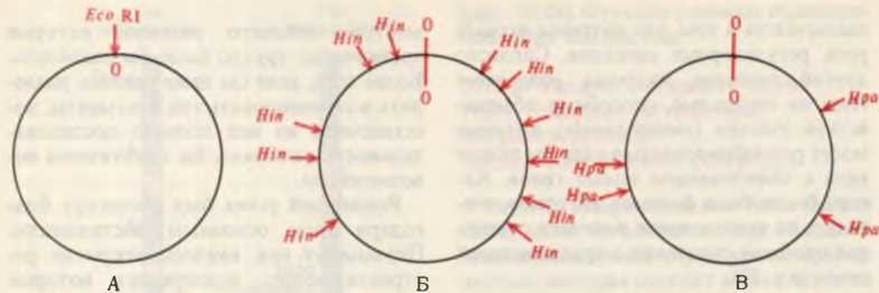
Рис. 27-31. Сайты рестрикции кольцевой молекулы ДНК SV40 для трех разных рестриктирующих эндонуклеаз - Eco RI, Hin и Нра I, каждая из которых узнает и катализирует расщепление двойной спирали в определенных участках. А. Eco RI расщепляет обе цепи кольцевой ДНК SV40 только в одном сайте, переводя ее в линейную форму. Этот сайт принят за точку отсчета. От него определяют положение сайтов рестрикции ДНК SV40 для других рестриктирующих эндонуклеаз. Б. Hin (смесь Hin dII и Hin dIII) расщепляет ДНК в одиннадцати сайтах с образованием двенадцати фрагментов. В. Нра I расщепляет ДНК всего в четырех местах с образованием пяти фрагментов. Данные ферменты были первыми рестриктирующими эндонуклеазами, которые Даниэль Натане и его коллеги применили для картирования генов в ДНК SV40. Сейчас уже известны сайты рестрикции генома SV40 для множества других эндонуклеаз.
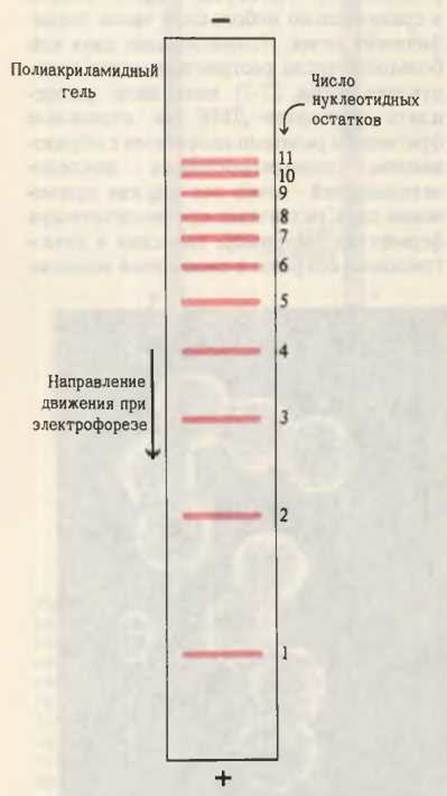
Рис. 27-32. Электрофоретическое разделение олигонуклеотидов по длине цепи на пластинке полиакриламидного геля. Чем короче олигонуклеотиды, тем быстрее они движутся к положительному электроду. Изменяя пористость полиакриламидного геля, данную процедуру можно использовать для разделения довольно длинных олигонуклеотидов, содержащих до двухсот и более остатков, даже если эти олигонуклеотиды различаются всего лишь одним остатком.
Вторым важным достижением оказалось усовершенствование электрофоретических методов разделения фрагментов ДНК в соответствии с числом содержащихся в них нуклеотидных остатков. Эти методы обладают настолько высоким разрешением, что позволяют разделять фрагменты ДНК размером до 200 нуклеотидов, даже если они отличаются друг от друга по длине всего лишь на один нуклеотид (рис. 27-32).
Третьим достижением стала разработка методов клонирования ДНК (гл. 30), которые сделали возможным получение достаточно больших количеств чистых генов - исходного материала для секвенирования. Было предложено два принципиальных подхода к секвенированию ДНК, у каждого из которых есть ряд вариантов. Фредерик Сэнгер, первым определивший аминокислотную последовательность белка, а именно инсулина (разд. 6.8), стал также первым ученым, установившим нуклеотидную последовательность в молекуле ДНК (бактериофага фХ174); произошло это в 1977 г. Сэнгер и его коллеги разработали очень изящную процедуру - метод терминации цепей, или, как его еще называют, “плюс-минус”-систему. Независимо от них Алан Максам и Уолтер Гилберт из США предложили несколько другой подход, получивший название химического метода. В обоих подходах используются фрагменты, полученные при расщеплении исходной ДНК с помощью рестриктирующих эндонуклеаз. В дополнении 27-1 описаны принципы метода определения нуклеотидной последовательности, разработанного Максамом и Гилбертом.
Дополнение 27-1. Секвенирование короткого фрагмента ДНК при помощи химического метода Максама-Гилберта
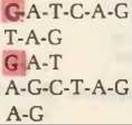
В приведенном ниже описании опущены некоторые детали для того, чтобы сконцентрировать внимание на основном принципе метода. Предположим, что у нас есть полученный в результате действия рестриктирующей эндонуклеазы фрагмент ДНК размером в 10 нуклеотидных остатков с последовательностью
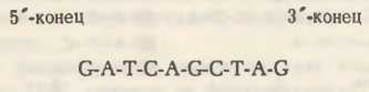
Первый этап состоит во введении радиоактивной метки в 5'-концевой остаток (G), который показан на красном фоне:
![]()
После этого меченный по 5'-положению олигонуклеотид распределяют на четыре порции. Первую из них подвергают химической обработке, в результате которой олигонуклеотид распадается на небольшие куски за счет статистического выщепления остатков С. При таком расщеплении олигонуклеотида по С может получиться следующая смесь:
В этом наборе фрагментов меченый 5'-концевой остаток показан на красном фоне. Обратите внимание на то, что два фрагмента помечены, а это значит, что они содержат 5'-конец исходного олигонуклеотида, в то время как фрагменты Т—А—G, А—G—С—Т—А—G и А—G не содержат метки, т. е. в них отсутствует исходный 5'-конец. Нас будут интересовать только меченые фрагменты.
Вторую порцию исходного меченого олигонуклеотида подвергают другой химической обработке, в результате которой выщепляются только остатки G, что приводит к образованию другого набора меченых фрагментов (рис. 1). Такую же обработку претерпевает и третья порция исходного меченого олигонуклеотида, который фрагментируется в результате выщепления только остатков А. Аналогичным образом, четвертую порцию расщепляют благодаря удалению только остатков Т. В конце концов имеют четыре различные смеси меченых фрагментов, полученные с помощью четырех разных химических процедур (рис. 1).
Каждую из четырех смесей фрагментов подвергают электрофорезу на пластинке геля в условиях, обеспечивающих разделение фрагментов в соответствии с числом содержащихся в них нуклеотидных остатков независимо от того, какие это остатки. При таком разделении фрагменты будут двигаться тем быстрее, чем они меньше. Точное положение каждого меченого фрагмента в геле определяют радиоавтографией. Положения меченых фрагментов в каждом из вариантов расщепления показаны на рис. 1. Положения немеченых фрагментов при этом не обнаруживаются, но эти фрагменты и не нужны для расшифровки последовательности.
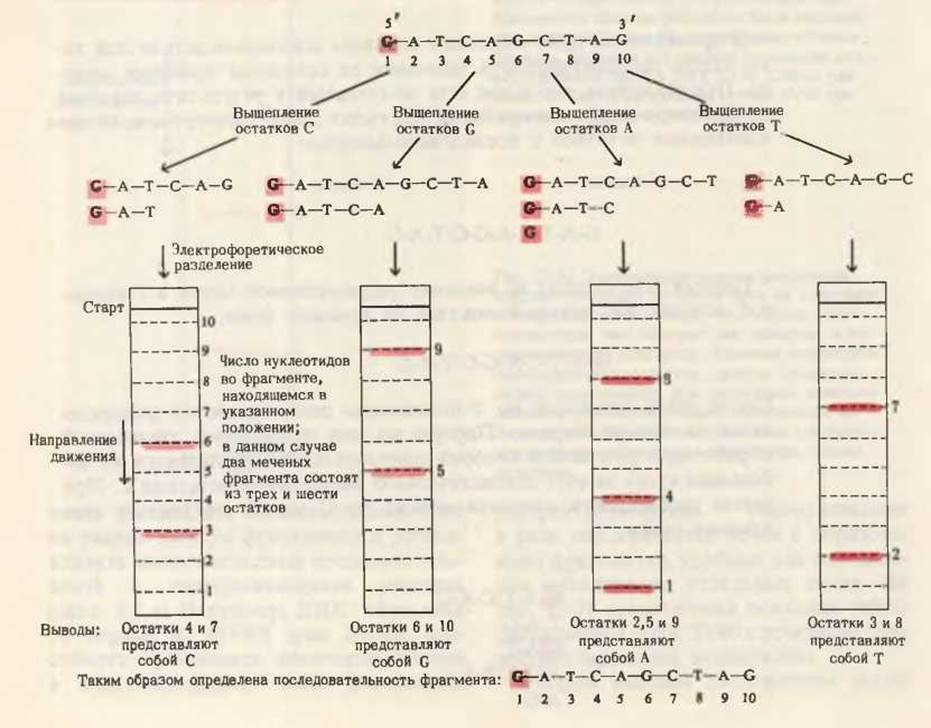
На рис. 1 результаты первого способа фрагментации, в котором выщеплялись остатки С, сопоставлены с электрофоретической картиной, показывающей расположение полученных при этом способе фрагментов длиной от 1 до 10 остатков. Здесь было обнаружено два меченых фрагмента. Очевидно, это именно те фрагменты, которые содержат 5'-конец исходного олигонуклеотида. Меченые фрагменты найдены в положениях, соответствующих олигонуклеотидам длиной в три и шесть остатков. Ясно, что в исходном нуклеотиде после этих меченых фрагментов в направлении к 3'-концу стоял остаток С, поскольку в химической процедуре, приведшей к расщеплению исходного олигонуклеотида, выщеплялся только этот остаток. Таким образом, в результате первой химической обработки мы узнали, что в положениях 4 и 7 (отсчитывая с 5'-конца) исходного олигонуклеотида должны располагаться остатки С.
Теперь точно так же поступают с тремя другими наборами фрагментов, полученными в результате специфического выщепления соответственно остатков G, А и Т из исходного олигонуклеотида (рис. 1). На схеме мы видим, что меченые фрагменты, полученные при выщеплении остатков G, движутся со скоростью, соответствующей олигонуклеотидам длиной в 5 и 9 нуклеотидных звеньев; таким образом, остатки 6 и 10 в исходном олигонуклеотиде должны быть G. Из третьего набора фрагментов, полученного удалением А, следует, что в положениях 2, 5 и 9 были остатки А; четвертый набор фрагментов, полученный выщеплением остатка Т, указывает на присутствие остатков Т в положениях 3 и 8. В нижней части рис. 1 дана нуклеотидная последовательность, установленная с помощью этой простой процедуры и некоторых рассуждений. Этим методом часто быстрее чем за 2 дня может быть определена нуклеотидная последовательность олигонуклеотидов, содержащих 200 и даже большее число остатков.
Чтобы расшифровать нуклеотидную последовательность целой молекулы ДНК, сначала ее фрагментируют с помощью рестриктирующей эндонуклеазы. Затем каждый из образовавшихся фрагментов секвенируют по отдельности по схеме, приведенной на рис. 1. Во второй аликвоте исходную ДНК расщепляют в других местах, используя другую рестриктирующую эндонуклеазу, и получают второй набор фрагментов. После того как секвенирование всех фрагментов второго набора завершено, сравнение двух наборов дает возможность найти участки перекрывания, необходимые для сборки фрагментов первого набора в правильном порядке. В результате может быть установлена нуклеотидная последовательность интересующей нас природной ДНК. Иногда, для того чтобы устранить неясности в некоторых участках последовательности, оставшиеся после первых двух расщеплений, приходится анализировать третий или четвертый наборы фрагментов.