Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Репликация и транскрипция ДНК
Матричная РНК синтезируется при помощи ДНК-зависимой РНК-полимеразы
Открытие ДНК-полимеразы и зависимости ее функционирования от присутствия ДНК-матрицы естественно привело к поиску ферментов, которые могли бы обеспечить синтез цепи РНК, комплементарной матрице ДНК. В 1959 г. такой фермент, способный образовывать полимер РНК из рибонуклеозид-5'-трифосфатов, был выделен из бактериальных экстрактов почти одновременно и независимо четырьмя разными группами американских биохимиков. Этот фермент, получивший название ДНК-зависимой РНК-полимеразы, по ряду свойств похож на ДНК-зависимую ДНК-полимеразу. РНК-полимеразе необходимы все четыре рибонуклеозид-5'-трифосфата (ATP, GTP, UTP и СТР) в качестве предшественников нуклеотидных элементов РНК, а также ионы Mg2+. В качестве существенного компонента активного центра она содержит цинк. РНК-полимераза удлиняет цепь РНК, присоединяя рибонуклеотидные мономеры к 3'-гидроксильному концу молекулы РНК, т.е. строит цепи РНК в направлении 5'→3'. Из ß- и у-фосфатных групп предшественников нуклеозид-5'-трифосфатов образуется неорганический пирофосфат. Удлинение цепи РНК описывается уравнением

РНК-полимераза требует для своего функционирования присутствия ДНК. Фермент наиболее активен, если в качестве матрицы используется природная двухцепочечная ДНК из того же самого или из какого-либо другого организма. Из двух цепей ДНК транскрибируется только одна. В места, соответствующие остаткам аденина ДНК-матрицы, в новосинтезированную РНК включаются остатки урацила (U), поскольку аденин и урацил образуют комплементарную пару. В места же, соответствующие тиминовым остаткам ДНК-матрицы, включаются остатки аденина. Остатки гуанина и цитозина в ДНК-матрице определяют включение соответственно цитозиновых и гуаниновых остатков в цепь РНК. Анализ нуклеотидного состава и нуклеотидной последовательности новообразованной РНК показал, что она обладает противоположной по отношению к матрице полярностью и её последовательность комплементарна последовательности матричной цепи. Хотя для работы РНК-полимеразы затравка не требуется, этот фермент не начинает функционировать до тех пор, пока не свяжется с особым участком матричной цепи ДНК, который служит сигналом инициации транскрипции. После связывания с этим участком фермент начинает синтез новой молекулы РНК; на ее 5'-конце обычно находится остаток GTP или АТР, чья 5'-трифосфатная группировка (обозначаемая ррр) не расщепляется до РРі и остается без изменений на протяжении всего процесса транскрипции. В ходе транскрипции новосинтезируемая цепь РНК временно образует (за счет спаривания ее оснований с основаниями матричной цепи ДНК) короткие отрезки гибридной двойной спирали ДНК-РНК, которые необходимы для правильного считывания цепи ДНК. Гибридный дуплекс существует лишь непродолжительное время, поскольку вскоре после синтеза РНК “сходит” с ДНК (рис. 28-17).
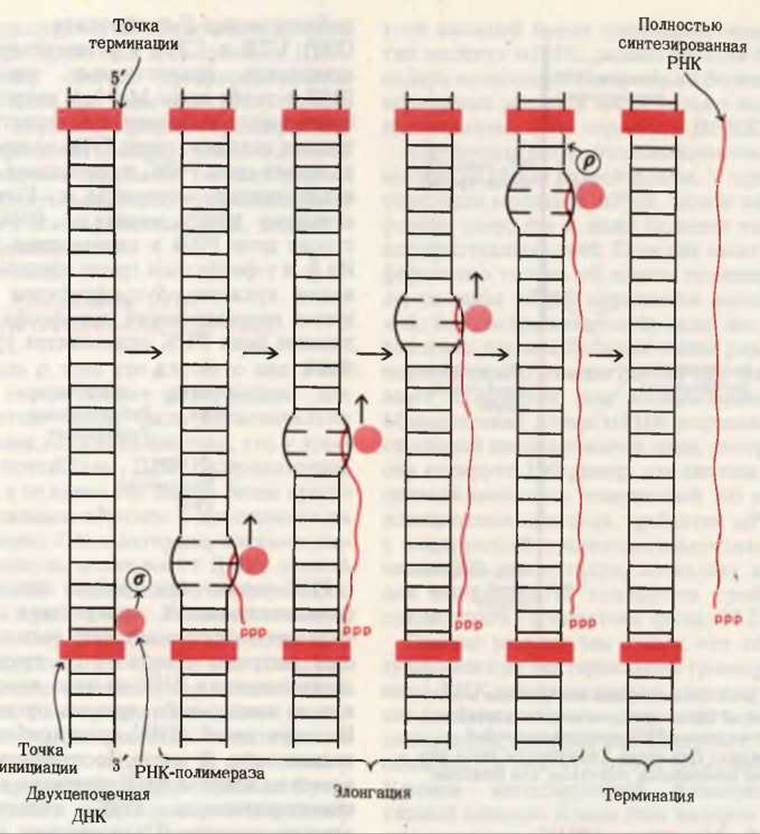
Рис. 28-17. Обобщенное изображение этапов транскрипции. РНК-полимераза прокариот должна сначала связаться с промоторным участком ДНК (гл. 29). Затем она движется к месту инициации синтеза РНК, где начинает транскрипцию со строго определенного нуклеотида, строя молекулу РНК в направлении 5'→3'. Приведенная схема не отражает действительной длины гибрида РНК-ДНК, которая составляет приблизительно 10 нуклеотидных пар. После инициации транскрипции субъединица σ отделяется от фермента. Для терминации синтеза цепи РНК необходима субъединица Р.
В Е. coli присутствует только одна ДНК-зависимая РНК-полимераза, которая способна синтезировать не только мРНК, но также тРНК и рРНК. Она представляет собой большой (мол. масса 500 000) и сложный фермент, состоящий из пяти полипептидных субъединиц: двух a-цепей, одной ß-, одной ß'- и одной σ-цепи. Структура холофермента этой полимеразы обозначается так: a2ßß'σ. Первый этап транскрипции - это присоединение холофермента к особому участку ДНК, называемому промотором, который представляет собой короткую последовательность, узнаваемую РНК-полимеразой. Разные промоторы несколько отличаются друг от друга по последовательности, что и определяет, вероятно, эффективность транскрипции разных генов. Как только РНК-полимераза заняла правильное положение в промоторном участке и образовала несколько фосфодиэфирных связей, субъединица σ отделяется от холофермента. Оставшийся “кор-фермент” (от англ. core - сердцевина) продолжает шаг за шагом удлинять молекулу РНК. Об окончании транскрибируемого гена (или генов) сигнализирует особая терминирующая последовательность в матрице ДНК. Для прекращения транскрипции и отделения РНК-полимеразы от ДНК необходим еще один специфический белок, обозначаемый р. Таким образом, синтез РНК включает три этапа: инициацию, элонгацию и терминацию (рис. 28-17).
Идентификации нуклеотидных последовательностей ДНК, в которых РНК-полимераза начинает и заканчивает транскрипцию, посвящены многочисленные исследования, однако пока еще многое относительно этих важных сигналов остается невыясненным.