Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
ДНК: структура хромосом и генов
Бактериальная ДНК защищена с помощью систем рестрикции-модификации
Уже давно было известно, что в бактериальной ДНК среди миллионов обычных оснований (А, Т, G и С) встречаются основания, несущие дополнительные метальные группы. Биологическое значение этих метилированных оснований стало понятным в результате ряда важных открытий, которые оказали большое влияние на развитие генетики и, в частности, биохимической генетики. Для каждого вида бактерий характерна своя особая картина распределения метилированных оснований по ДНК, отличающая ее от ДНК других видов. Если ДНК какого-либо другого вида каким-то образом проникнет в живую бактериальную клетку, то она будет признана там “чужеродной” именно по отсутствию в ней специфической для данного вида картины распределения метилированных оснований, присущей ДНК клеток этого вида. В такой ситуации чужеродная ДНК будет разрушена специфической нуклеазой, которая расщепляет обе цепи ДНК непосредственно в том месте, где отсутствуют характерные для ДНК клетки-хозяина метилированные основания, или вблизи этого места. Таким образом чужеродные ДНК подвергаются рестрикции: они разрушаются с помощью специфических нуклеаз, вырабатываемых каждым видом бактерий.
ДНК бактерий данного вида защищена двумя близкими по своей специфичности ферментами: 1) модифицирующей метилазой и 2) рестриктирующей эндонуклеазой. Модифицирующая метилаза отвечает за образование специфической для данного вида картины метилирования в определенных коротких последовательностях собственной ДНК клетки. Метальные группы в этих последовательностях остаются без изменений в течение всей жизни клетки. Соответствующая рестриктирующая эндонуклеаза в свою очередь расщепляет обе цепи любой другой ДНК, в которой эта специфические последовательности оснований не метилированы. Примером может служить рестриктирующая эндонуклеаза HindII бактерии Haemophilus influenzae (каждая рестриктирующая эндонуклеаза имеет свое обозначение). Этот фермент расщепляет обе цепи любой ДНК, содержащей определенную последовательность оснований в местах, указанных стрелками:

но не расщепляет эту же последовательность, если основания, отмеченные красной звездочкой, метилированы:
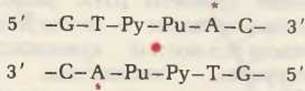
Интересно, что этот короткий участок ДНК, метилированный или неметилированный, обладает внутренней симметрией относительно центральной точки, показанной красным цветом. Если повернуть этот участок на 180° в плоскости рисунка вокруг центральной точки, то он будет читаться точно так же, как до поворота. Такой тип симметрии характеризуется осью симметрии второго порядка. Большинство проверенных последовательностей модификации-рестрикции обладают симметрией второго порядка. Рестриктирующая эндонуклеаза Hind II расщепляет обе цепи в середине этого участка в любой ДНК, в которой эта последовательность не метилирована. Будучи расщепленной таким образом, чужеродная ДНК не может быть исправлена и поэтому не может реплицироваться.
В дальнейшем под действием других клеточных нуклеаз такая ДНК разрушается до мононуклеотидов.
В табл. 27-7 приведены специфические последовательности, атакуемые типичными рестриктирующими эндонуклеазами (обозначенными символами) из разных видов бактерий. Каждая из узнаваемых таким ферментом последовательностей обладает симметрией второго порядка. В зависимости от типа рестриктирующей эндонуклеазы в месте расщепления двухцепочечной ДНК образуются либо “тупые”, т. е. заполненные концы (как в случае описанной выше Hind II), либо выступающие концы (примером может служить эндонуклеаза Eco RI из Е. coli). В последнем случае два перекрывающихся конца называют липкими, поскольку они способны образовать друг с другом комплементарные пары оснований.
Таблица 21-1. Специфичность некоторых рестриктирующих эндонуклеаз1)
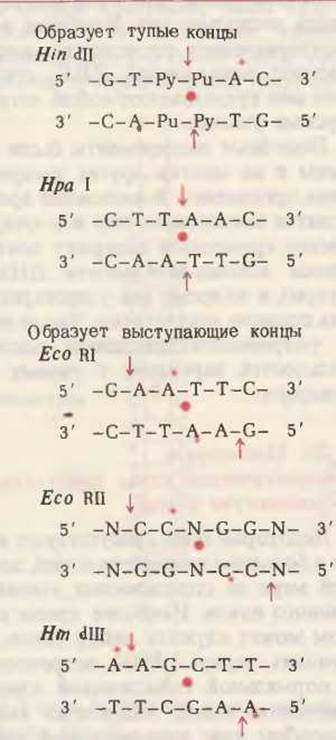
1) Жирной красной точкой обозначена ось симметрии второго порядка, красными стрелками показаны места расщепления. Красными звездочками указаны места метилирования (если они известны) в организме, из которого данный фермент выделен: Haemophilus influenza для Hin dH и Hin dill, E. coli для Eco RI и Eco RII и Haemophilus parainfluenza для Hpa II. Pu - пурин, Ру - пиримидин, N - A или T.
Было подсчитано, что последовательности из шести нуклеотидов с осевой симметрией второго порядка могут встречаться в любой ДНК независимо от вида с вероятностью 1 : 4000. Поскольку молекулы ДНК бактерий состоят из миллионов нуклеотидных пар, вероятность того, что любая бактериальная ДНК расщепится хотя бы один раз любой данной рестриктирующей эндонуклеазой, очень велика. Однако клетка-хозяин защищает свою собственную ДНК путем метилирования одного или нескольких оснований в последовательности, подверженной рестрикции; метилированная последовательность не связывается с рестриктирующей эндонуклеазой и потому не расщепляется.
У разных видов бактерий обнаружено уже свыше 150 различных рестриктирующих эндонуклеаз. Некоторые бактерии содержат больше одного набора модифицирующих метилаз и рестриктирующих эндонуклеаз. Однако ДНК вирусов бактерий научились с помощью ряда способов преодолевать рестрикционную защиту клеток их хозяев. Некоторые вирусные ДНК содержат разного рода модифицированные основания, которые позволяют им избегать расщепления рестриктирующими эндонуклеазами клетки-хозяина, в которую они попали. Модифицирующими группами в таких вирусных ДНК служат метальные, гидроксиметильные и глюкозильные группы. Другие вирусы в ходе эволюции приобрели такие последовательности в ДНК, которые не содержат участков, узнаваемых некоторыми рестриктирующими эндонуклеазами.
Рестриктирующие нуклеазы оказались исключительно полезными в качестве инструмента в генетических исследованиях, поскольку они позволяют воспроизводимо осуществлять расщепление обеих цепей ДНК в строго определенных точках. Обнаружение ферментов с такими свойствами открыло новую эру в биохимии генов. Благодаря рестриктирующим эндонуклеазам, многие из которых в настоящее время производятся фирмами и поступают в продажу, стало возможным целенаправленное разрезание и картирование хромосом; эти ферменты стали также необходимым инструментом при определении нуклеотидных последовательностей ДНК. Рестриктирующие эндонуклеазы позволили начать работы по встраиванию генов одного организма в геном другого (гл. 30). Ученым, открывшим явление рестрикции ДНК, изучившим природу действия рестриктирующих эндонуклеаз и показавшим возможность их использования для вырезания генов, - Вернеру Арберу из Швейцарии, Гамильтону Смиту и Даниэлю Натансу (оба последних из США) в 1978 г. была присуждена Нобелевская премия по физиологии и медицине.