Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Гормоны мозгового вещества надпочечников
Биосинтез катехоламинов
Катехоламиновые гормоны — дофамин, норадреналин и адреналин — представляют собой 3,4-дигидроксипроизводные фенилэтиламина. Они синтезируются в хромаффинных клетках мозгового слоя надпочечников. Свое название эти клетки получили потому, что содержат гранулы, окрашивающиеся под действием бихромата калия в краснокоричневый цвет. Скопления таких клеток обнаружены также в сердце, печени, почках, половых железах, адренергических нейронах постганглионарной симпатической системы и в центральной нервной системе.
Главный продукт мозгового слоя надпочечников — адреналин. На долю этого соединения приходится примерно 80% всех катехоламинов мозгового слоя. Вне мозгового вещества адреналин не образуется. В отличие от него норадреналин, обнаруживаемый в органах, иннервируемых симпатическими нервами, образуется преимущественно in situ (~ 80% общего количества); остальная часть норадреналина также образуется главным образом в окончаниях нервов и достигает своих мишеней с кровью.
Превращение тирозина в адреналин включает четыре последовательных этапа: 1) гидроксилирование кольца, 2) декарбоксилирование, 3) гидроксилирование боковой цепи и 4) N-метилирование. Путь биосинтеза катехоламинов и участвующие в нем ферменты представлены на рис. 49.1 и 49.2.
Тирозин-гидроксилаза
Тирозин — непосредственный предшественник катехоламинов, а тирозин-гидроксилаза лимитирует скорость всего процесса биосинтеза катехоламинов. Этот фермент встречается как в свободном виде, так и в связанной с субклеточными частицами форме. С тетрагидроптеридином в качестве кофактора он выполняет оксидоредуктазную функцию, превращая L-тирозин в L-дигидроксифенилаланин (L-ДОФА). Существуют различные пути регуляции тирозин- гидроксилазы как скорость-лимитирующего фермента. Наиболее важный из них заключается в ингибировании катехоламинами по принципу обратной связи: катехоламины конкурируют с ферментом за птеридиновый кофактор, образуя с последним шиффово основание. Тирозин-гидроксилаза, кроме того, конкурентно ингибируется рядом производных тирозина, в том числе а-метилтирозином. В некоторых случаях это соединение используют для блокады избыточной продукции катехоламинов при феохромоцитоме, однако существуют более эффективные средства, обладающие к тому же менее выраженным побочным действием. Соединения еще одной группы подавляют активность тирозин-гидроксилазы, образуя комплексы с железом и удаляя таким путем имеющийся кофактор. В качестве примера такого соединения можно привести а, а'1-дипиридил.
Катехоламины не проникают через гематоэнцефалический барьер, и, следовательно, их присутствие в мозге должно объясняться местным синтезом. При некоторых заболеваниях центральной нервной системы, например болезни Паркинсона, наблюдаются нарушения синтеза дофамина именно в мозге. Предшественник дофамина — L-ДОФА — легко преодолевает гематоэнцефалический барьер и поэтому служит эффективным средством лечения болезни Паркинсона.
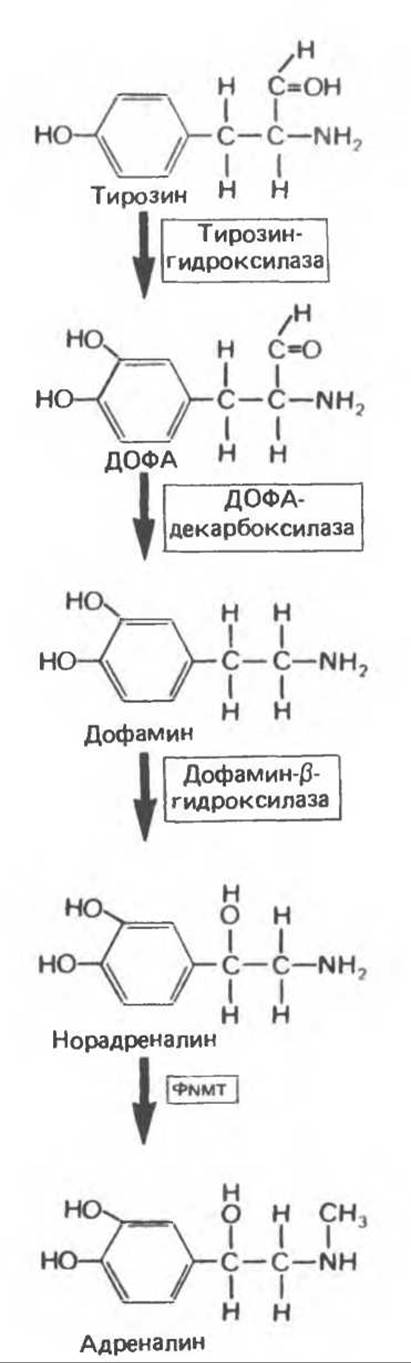
Рис. 49.1. Биосинтез катехоламинов. ФNMT — фенилэтаноламин-N-метилтрансфераза. (Modified and reproduced, with permission, from Goldfien A. The adrenal medulla. In: Basic and Clinical Endocrinology, 2nd ed. Greenspan FS, Forsham PH [editors]. Appleton and Lange, 1986.)
ДОФА-декарбоксилаза
В отличие от тирозин-гидроксилазы. обнаруживаемой лишь в тканях, способных синтезировать катехоламины, ДОФА-декарбоксилаза присутствует во всех тканях. Этому растворимому ферменту требуется пиридоксальфосфат для превращения L- ДОФА в 3,4-дигидроксифенилэтиламин (дофамин). Реакция конкурентно ингибируется соединениями, напоминающими L-ДОФА, например а-метил-ДОФА. Галоидзамещенные соединения образуют с L-ДОФА шиффово основание и также ингибируют реакцию декарбоксилирования.
а-Метил-ДОФА и другие родственные соединения, такие, как 3-гидрокситирамин (образующийся из тирамина), а-метилирозин и метараминол, с успехом используются для лечения некоторых форм гипертонии. Антигипертензивное действие этих метаболитов обусловлено, по-видимому, их способностью стимулировать а-адренергические рецепторы (см. ниже) кортикобульбарной системы в центральной нервной системе, что приводит к уменьшению активности периферических симпатических нервов и снижению артериального давления.
Дофамин-β-гидроксилаза
Дофамин-β-гидроксилаза (ДБГ) — оксидаза со смешанной функцией, катализирующая превращение дофамина в норадреналин. ДБГ использует аскорбат в качестве донора электронов, а фумарат — в качестве модулятора; в активном центре фермента содержится медь. ДБГ клеток мозгового слоя надпочечников локализуется, вероятно, в секреторных гранулах. Таким образом, превращение дофамина в норадреналин происходит в этих органеллах. ДБГ высвобождается из клеток мозгового слоя надпочечников и нервных окончаний вместе с норадреналином, но (в отличие от последнего) не подвергается обратному захвату нервными окончаниями.
Фенилэтаноламин — N-метилтрансфераза
Растворимый фермент фенилэтаноламин — N-метилтрансфераза (ФNМТ) катализирует N-метилирование норадреналина с образованием адреналина в адреналин-продуцирующих клетках мозгового слоя надпочечников. Поскольку данный фермент растворим, можно предположить, что превращение норадреналина в адреналин происходит в цитоплазме. Синтез ФNМТ стимулируется глюкокортикоидными гормонами, проникающими в мозговой слой по внутринадпочечниковой портальной системе. Эта система обеспечивает в 100 раз большую концентрацию стероидов в мозговом слое, чем в системной артериальной крови. Столь высокая их концентрация в надпочечниках, по-видимому, необходима для индукции ФNMT.