Биохимия человека Том 2 - Марри Р. 1993
Частные вопросы
Питание, пищеварение и всасывание
Панкреатическое и кишечное пищеварение
Желудочное содержимое, или химус, в ходе переваривания периодически поступает в двенадцатиперстную кишку через пилорический клапан. Панкреатический и желчный протоки открываются в двенадцатиперстную кишку в непосредственной близости к пилорусу. Щелочное содержимое секрета поджелудочной железы и желчи нейтрализует химус и сдвигает его pH в щелочную сторону. Этот сдвиг pH необходим для проявления активности ферментов панкреатического и кишечного сока, но он ингибирует дальнейшее действие пепсина.
Желчь
Кроме многочисленных функций в межуточном обмене, печень в силу ее способности продуцировать желчь играет важную роль в пищеварении. Желчный пузырь, мешотчатый орган, примыкающий к печеночному протоку, накапливает некоторое количество желчи, образуемой печенью в периоды между приемами пищи. Во время пищеварения желчный пузырь сокращается и быстро выбрасывает желчь в тонкий кишечник через общий желчный проток. Продукты секреции поджелудочной железы смешиваются с желчью, поскольку попадают в общий желчный проток чуть раньше его впадения в двенадцатиперстную кишку.
А. Состав желчи. Печеночная желчь отличается по составу от желчи, содержащейся в желчном пузыре. Последняя, как показано в табл. 53.10, является более концентрированной.
Б. Функции желчи.
1. Эмульсификация. Соли желчных кислот обладают способностью значительно уменьшать поверхностное натяжение. Благодаря этому они осуществляют эмульгирование жиров в кишечнике, растворяют жирные кислоты и нерастворимые в воде мыла. Присутствие желчи в кишечнике способствует завершению переваривания и всасывания жиров, а также всасыванию жирорастворимых витаминов A, D, Е и К. При нарушении переваривания жира плохо перевариваются также другие пищевые вещества, потому что жир обволакивает частицы пищи и препятствует действию на них ферментов. В этих условиях деятельность кишечных бактерий приводит к активации процессов гниения и образования газа.
2. Нейтрализация кислоты. Помимо своих пищеварительных функций желчь, имеющая pH чуть выше 7, нейтрализует кислый химус, поступающий из желудка, подготавливая его для переваривания в кишечнике.
Таблица 53.10. Состав печеночной и пузырной желчи
|
Печеночная желчь (секретируемая) |
Пузырная желчь |
||
|
Процент от цельной желчи |
Процент от общего количества твердых веществ |
Процент от цельной желчи |
|
|
Вода |
97,00 |
.... |
85,92 |
|
Твердые вещества |
2,52 |
.... |
14,08 |
|
Желчные кислоты |
1,93 |
36,9 |
9,14 |
|
Муцин и пигменты |
0,53 |
21,3 |
2,98 |
|
Холестерол |
0,06 |
2,4 |
0,26 |
|
Эстерифицированные и неэстерифицированные жирные кислоты |
0,14 |
5,6 |
0,32 |
|
Неорганические соли |
0,84 |
33,3 |
0,65 |
|
Удельный вес |
1,01 |
.... |
1,04 |
|
pH |
7,1—7,3 |
.... |
6.9—7,7 |
3. Экскреция. Желчь — важный носитель экскретируемых желчных кислот и холестерола, но она также удаляет из организма многие лекарства, токсины, желчные пигменты и различные неорганические вещества, такие, как медь, цинк и ртуть.
4. Растворимость холестерола в желчи; образование желчных камней. Свободный холестерол нерастворим в воде и, следовательно, включается в мицеллы, образуемые фосфатидилхолином и желчными солями. Более того, фосфатидилхолин, преобладающий фосфолипид желчи, сам по себе нерастворим в водных системах, но он может быть переведен в растворимое состояние желчными солями в составе мицелл. Большие количества холестерола, присутствующие в желчи человека, подвергаются солюбилизации в этих водорастворимых смешанных мицеллах, что обеспечивает перенос холестерола в кишечник через желчный проток. Однако фактическая растворимость холестерола в желчи зависит от соотношения желчных солей, фосфатидилхолина и холестерола. Она зависит также от содержания воды в желчи, что особенно важно в случае разбавленной печеночной желчи.
Используя систему треугольных координат (рис. 53.2), Редингер и Смолл сумели определить максимальную растворимость холестерола в желчи из желчного пузыря человека. Из чертежа видно, что любая точка в этой системе, находящаяся выше кривой АВС, будет представлять желчь такого состава, в которой холестерол окажется либо в состоянии перенасыщенного раствора, либо выпадет в осадок.
Можно полагать, что у больных желчнокаменной болезнью в какой-то период жизни образуется аномальная желчь, перенасыщенная холестеролом. С течением времени различные факторы, например инфекция, могут выступать в качестве агентов, провоцирующих выпадение из перенасыщенной желчи избытка холестерола в виде кристаллов. Если новообразованные кристаллы сразу не перенесутся с желчью в кишечник, то они будут расти, образуя камни. Определение активности ключевых ферментов образования желчных кислот в печени пациентов с желчнокаменной болезнью показало, что синтез холестерола у них повышен, а синтез желчных кислот снижен, в результате чего возрастает концентрация холестерола в печени. Снижение активности 7а-гидроксилазы может приводить к уменьшению энтерогепатического запаса желчных кислот, что служит для печени сигналом к образованию еще больших количеств холестерола. Желчь оказывается перегруженной холестеролом, который не может полностью раствориться в смешанных мицеллах.
Основываясь на изложенных выше данных о растворимости холестерола, была сделана попытка разработать способ растворения желчных камней или предотвращения их дальнейшего образования. Применение хенодезоксихолевой кислоты открывает возможность специфического лечения больных с рентгеноплотными камнями в функционирующем желчном пузыре, поскольку это соединение обладает способностью ингибировать гидроксиметилглутарил (ГМГ)-СоА-редуктазу в печени, что приводит к снижению синтеза холестерола.
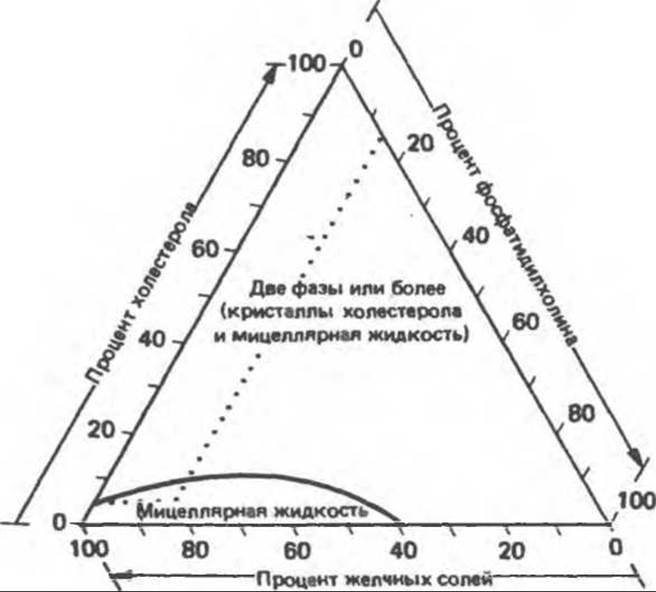
Рис. 53.2. Способ изображения трех основных компонентов желчи (желчные кислоты, фосфатидилхолин и холестерол) в треугольных координатах. Каждый компонент выражен в процентах общего количества (в молях) желчных солей, фосфатидилхолина и холестерола. Кривая отражает максимальную растворимость холестерола в различных смесях желчных солей и фосфатидилхолина. Точка пересечения пунктирных линий соответствует нормальной желчи (5% холестерола, 15% фосфатидилхолина и 80% желчных солей) и находится в зоне однофазной мицеллярной жидкости. Желчь, которая по своему составу располагается выше этой линии, должна содержать избыток холестерола в виде перенасыщенного раствора либо осадка (кристаллы или жидкие кристаллы). (Reproduced, with permission, from Redinger R. N. Small D. M. Bile composition, bile salt metabolism, and gallstones. Arch. Intern. Med., 1972, 130, 620. Copyright © 1972. American Medical Association.)
5. Метаболизм желчных пигментов. Происхождение желчных пигментов из гемоглобина обсуждается в гл. 33.
Переваривание секретом поджелудочной железы
Секрет поджелудочной железы — это неклейкая водянистая жидкость, сходная со слюной по количеству воды, содержащая белок, а также органические и неорганические ионы (преимущественно Na+, К+, НСО-3 и Сl-) и, кроме того, малые количества Са2+, Zn2+, НРO2-4 и SO2+4. Значение pH панкреатического секрета — отчетливо щелочное — 7,5—8,0 или выше.
В секрете присутствуют многие ферменты, некоторые из них секретируются в виде зимогенов.
A. Трипсин, химотрипсин и эластаза. Протеолитическое действие панкреатического секрета обусловливается тремя эндопептидазами — трипсином, химотрипсином и эластазой, которые расщепляют белки и полипептиды, поступающие из желудка, с образованием полипептидов, пептидов или тех и других. Трипсин специфически действует на пептидные связи, образуемые основными аминокислотами, химотрипсин — на связи между остатками незаряженных аминокислот (например, ароматических), в то время как эластаза вопреки своему названию обладает довольно широкой специфичностью, расщепляя связи, примыкающие к остаткам малых аминокислот, таких, как глицин, аланин и серин. Все три фермента секретируются в виде зимогенов. Активация трипсиногена осуществляется другим протеолитическим ферментом, энтерокиназой, секретируемой слизистой кишечника. Она гидролизует лизиновую пептидную связь в зимогене, высвобождая малый полипептид, что приводит к разворачиванию молекулы в активный трипсин. Образовавшийся трипсин действует не только на новые молекулы трипсиногена, но и на другие зимогены панкреатического секрета — химотрипсиноген, проэластазу и прокарбоксипептидазу — с высвобождением соответственно химотрипсина, эластазы и карбоксипептидазы.
Б. Карбоксипеотидаза. Дальнейшее расщепление полипептидов, образовавшихся под действием эндопептидаз, осуществляет экзопептидаза — карбоксипептидаза, которая атакует С-концевую пептидную связь, высвобождая одиночные аминокислоты.
B. Амилаза. Крахмал-расщепляющая активность секрета поджелудочной железы обусловлена панкреатической а-амилазой. Она сходна по действию с амилазой слюны и гидролизует крахмал и гликоген с образованием мальтозы, мальтотриозы [три а-глюкозных остатка, связанные а(1→4)-связями], a также смеси разветвленных (1→6) олигосахаридов (а-декстрины), неразветвленных олигосахаридов и некоторого количества глюкозы.
Г. Липаза. Панкреатическая липаза действует на поверхности раздела жир — вода тонкоэмульгированных липидных капель, образуемых в кишечнике при механическом перемешивании в присутствии продуктов действия липазы языка, желчных солей, колнпазы (белка, присутствующего в секрете поджелудочной железы), фосфолипидов и фосфолипазы А2 (также входящей в состав панкреатического секрета). Фосфолипаза А2 и колипаза секретируются в виде про-форм, и для их активации требуется триптический гидролиз специфических пептидных связей. Для проявления активности фосфолипазы А2 необходим Са2+. В результате ограниченного гидролиза фосфолипазой А2 эфирной связи фосфолипидов в положении 2 (см. рис. 25.5) липаза связывается на поверхнсти раздела субстрата и происходит быстрый гидролиз триацилглицеролов. Колипаза связывается с поверхностью раздела системы желчная соль — триацилглицерол/вода, образуя высокоаффинный якорь для липазы. Полный гидролиз триацилглицеролов приводит к образованию глицерола и жирных кислот. Заметим, однако, что отщепление второй и третьей жирных кислот от триацилглицеролов происходит с возрастающей трудностью. Панкреатическая липаза в сущности специфична в отношении гидролиза первичных эфирных связей, т. е. связей в положениях 1 и 3 триацилглицеролов. Во время переваривания жира водная, или «мицеллярная», фаза содержит смешанные дисковидные мицеллы и липосомы из желчных солей, насыщенных продуктами липолиза (см. рис. 15.34). Так как гидролиз вторичной эфирной связи в триацилглицероле затруднен, можно предположить, что перевариванию триацилглицерола предшествует удаление терминальных жирных кислот с образованием 2-моноацилглицерола. Поскольку последняя жирная кислота связана вторичной эфирной связью, ее удаление требует изомеризации в первичную эфирную связь. Это относительно медленный процесс, и поэтому главными конечными продуктами переваривания триацилглицерола оказываются именно 2-моноацилглицеролы и только менее одной четверти переваренного триацилглицерола полностью распадается на глицерол и жирные кислоты (рис. 53.3).
Д. Гидролаза холестериловых эфиров (холестеролэстераза). В условиях, характерных для просвета кишечника, фермент катализирует гидролиз эфиров холестерола, который затем всасывается из кишечника в неэстерифицированной, свободной форме.
Е. Рибонуклеаза (РНКаза) и дезоксирибонуклеаза (ДНКаза) получены из ткани поджелудочной железы (см. гл. 38 и 39).
Ж. Фосфолипаза А2. Фосфолипаза А2 гидролизует эфирную связь во 2-м положении глицерофосфолипидов как желчного, так и пищевого происхождения с образованием лизофосфолипидов.
Переваривание секретом кишечника
Кишечный сок, секретируемый железами Бруннера и Либеркюна, также содержит пищеварительные ферменты, в число которых входят:
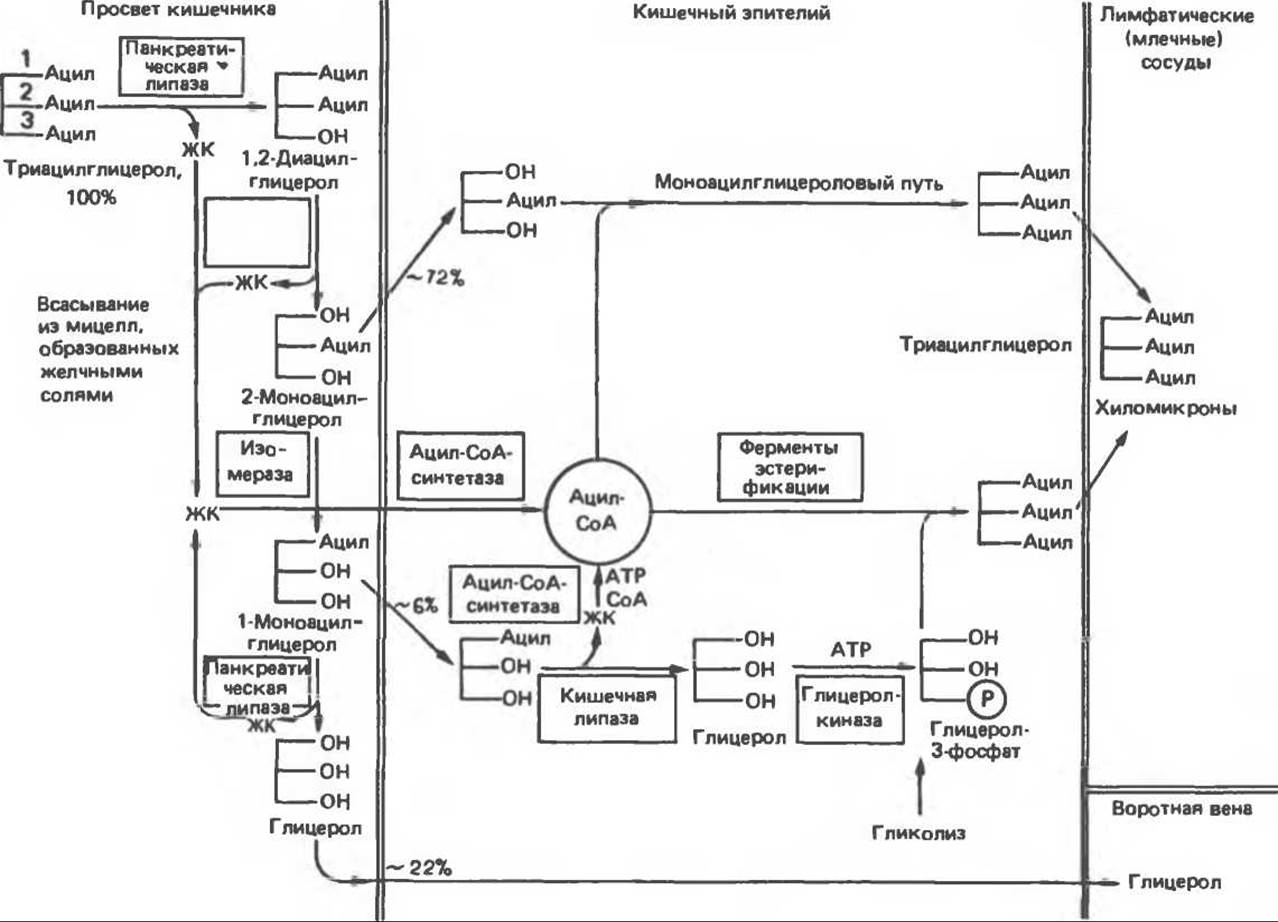
Рис. 53.3. Переваривание и всасывание триацилглицеролов. ЖК — длинноцепочечные жирные кислоты. (Modified from Mattson F. Н., Volpenheim R.A. The digestion and absorption of triglycerides. J. Biol. Chem., 1964, 239, 2772.)
1) аминопептидаза, представляющая собой экзопептидазу, которая гидролизует пептидные связи за N-концевыми аминокислотами полипептидов и олигопептидов; дипептидазы различной специфичности, некоторые из них могут находиться внутри кишечного эпителия; они завершают расщепление дипептидов до свободных аминокислот;
2) специфические дисахаридазы и олигосахаридазы, такие, как а-глюкозидаза (мальтаза), удаляющая единичные глюкозные остатки из а(1→4)-связанных олигосахаридов и дисахаридов начиная с нередуцирующих концов, изомальтаза (а-декстриназа), которая гидролизует (1→6)-связи а-декстринов; ß- галактозидаза (лактаза), удаляющая галактозу из лактозы; сахараза, гидролизующая сахарозу, и трегалаза. расщепляющая трегалозу;
3) фосфатаза, удаляющая фосфат из некоторых органических фосфатов (гексозофосфаты и глицерофосфат) и из нуклеотидов пищевого происхождения или образующихся из нуклеиновых кислот в результате их переваривания нуклеазами;
4) полинуклеотидазы, которые расщепляют нуклеиновые кислоты на нуклеотиды;
5) нуклеозидазы (нуклеозидфосфорилазы), катализирующие фосфоролиз нуклеозидов с образованием свободных азотистых оснований и пентозофосфатов;
6) кишечный секрет, по-видимому, содержит также фосфолипазу, которая действует на фосфолипиды с образованием глицерола, жирных кислот, фосфорной кислоты и оснований, таких, как холин.
Основные продукты переваривания
Окончательный результат действия описанных пищеварительных ферментов заключается в редуцировании компонентов пищи до форм, которые могут всасываться и усваиваться. Этими конечными продуктами переваривания являются для углеводов моносахариды (в основном глюкоза), для белков — аминокислоты, для триацилглицеролов — жирные кислоты, глицерол и моноацилглицерол и для нуклеиновых кислот — основания, нуклеозиды и пентозы.
Полисахариды стенки растительной клетки и лигнин, которые не расщепляются ферментами млекопитающих, составляют пищевые волокна и образуют массу, остающуюся после переваривания. Волокна выполняют важную функцию, придавая пище дополнительный объем, что уже обсуждалось в этой главе. Основные пищеварительные процессы суммированы в табл. 53.11.