Биохимия человека Том 2 - Марри Р. 1993
Структура, функция и репликация информационных макромолекул
Технология рекомбинантных ДНК
РНК
Химическая природа РНК
Рибонуклеиновая кислота представляет собой сополимер пуриновых и пиримидиновых рибонуклеотидов, соединенных друг с другом, как и в ДНК, 3'—5'-фосфодиэфирными мостиками (рис. 37.6). Хотя эти два вида нуклеиновых кислот имеют много общего, по ряду признаков они отличаются друг от друга.
1. У РНК углеводным остатком, к которому присоединены пуриновые или пиримидиновые основания и фосфатные группы, является рибоза, а не 2'-дезоксирибоза (как у ДНК).
2. Пиримидиновые компоненты РНК отличаются от таковых у ДНК. В состав РНК, как и в состав ДНК, входят нуклеотиды аденина, гуанина и цитозина. В то же время РНК (за исключением некоторых специальных случаев, на которых мы остановимся ниже) не содержит тимина, его место в молекуле РНК занимает урацил.
3. РНК — одноцепочечная молекула (в отличие от ДНК, имеющей двухцепочечную структуру), однако при наличии в цепи РНК участков с комплементарной последовательностью (противоположной полярности) единичная цепь РНК способна сворачиваться с образованием так называемых «шпилек», структур, имеющих двухспиральные характеристики (рис. 37.7).
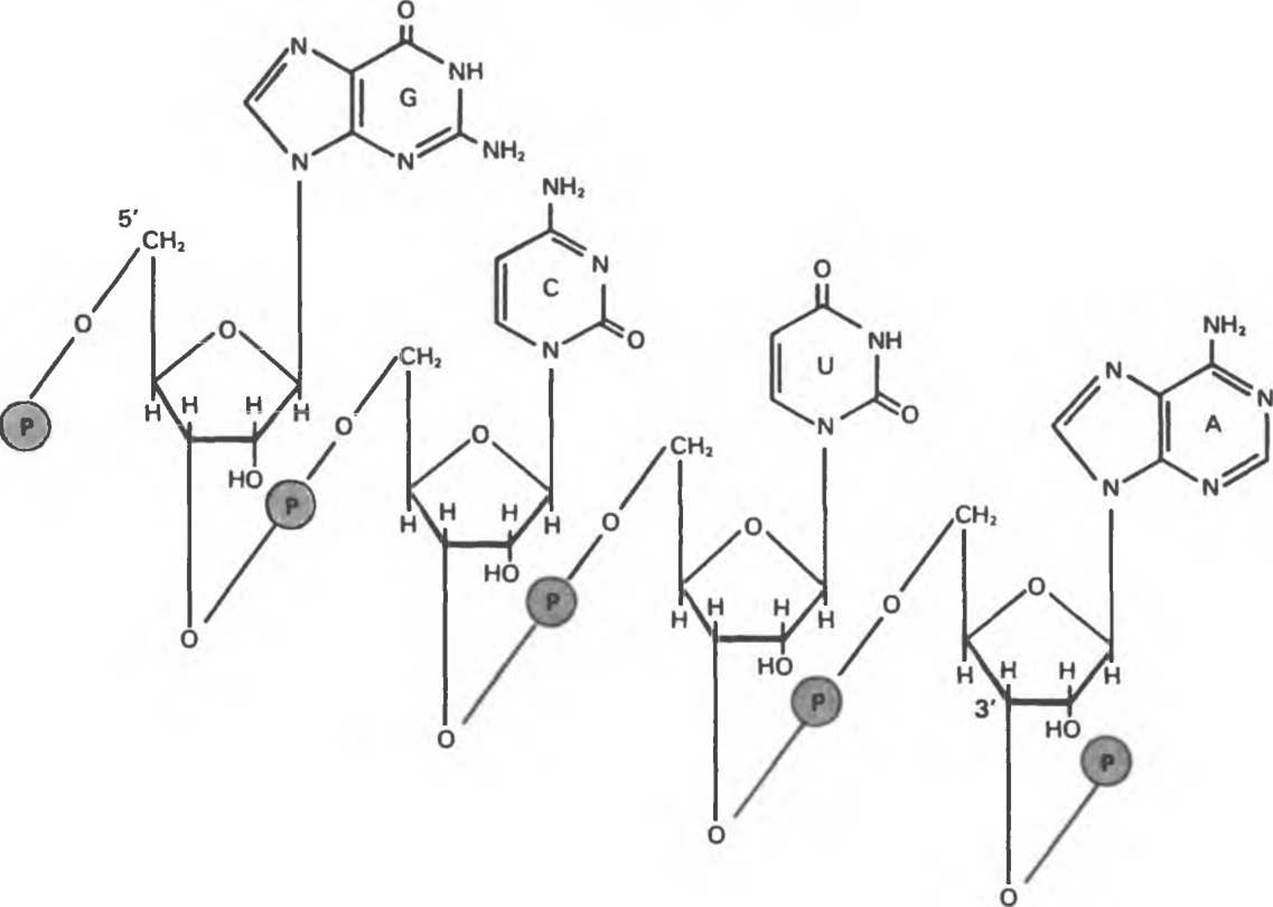
Рис. 37.6. Фрагмент молекулы рибонуклеиновой кислоты (РНК), в котором пуриновые и пиримидиновые основания — аденин (А), урацил (U), цитозин (С) и гуанин (G)—удерживаются фосфодиэфирным остовом, соединяющим рибозильные остатки, связанные N-гликозидной связью с соответствующими нуклеиновыми основаниями. Обратите внимание: цепь РНК обладает определенной направленностью, на которую указывают 5'- и 3'-концевые фосфатные остатки.
4. Так как молекула РНК представляет собой одиночную цепь, комплементарную только одной из цепей ДНК, содержание в ней гуанина не обязательно равно содержанию цитозина, а содержание аденина не обязательно равно содержанию урацила.
5. РНК может быть гидролизована щелочью до 2', 3'-циклических диэфиров мононуклеотидов; в роли промежуточного продукта гидролиза выступает 2', 3', 5'-триэфир, который не образуется при щелочном гидролизе ДНК из-за отсутствия у последней 2'-гидроксильных групп; щелочная лабильность РНК (сравнительно с ДНК) является полезным свойством как для диагностических, так и для аналитических целей.
Информация, содержащаяся в одноцепочечной РНК, реализуется в виде определенной последовательности пуриновых и пиримидиновых оснований (т. е. в первичной структуре) полимерной цепи. Эта последовательность комплементарна кодирующей цепи гена, с которой «считывается» РНК. Вследствие комплементарности молекула РНК способна специфически связываться (гибридизоваться) с кодирующей цепью, но не гибридизуется с некодирующей цепью ДНК. Последовательность РНК (за исключением замены Т на U) идентична последовательности некодирующей цепи гена (рис. 37.8).
Биологические функции РНК
Известно несколько видов РНК. Почти все они непосредственно вовлечены в процесс биосинтеза белка. Молекулы цитоплазматической РНК, выполняющие функции матриц белкового синтеза, называются матричными РНК (мРНК). Другой вид цитоплазматической РНК — рибосомная РНК (рРНК) — выполняет роль структурных компонентов рибосом (органелл, играющих важную роль в синтезе белка). Адапторные молекулы транспортных РНК (тРНК) участвуют в трансляции (переводе) информации мРНК в последовательность аминокислот в белках.
Значительная часть РНК — первичных транскриптов, образующихся в эукариотических клетках, включая и клетки млекопитающих, — подвергается деградации в ядре и не играет какой-либо структурной или информационной роли в цитоплазме. В культивируемых клетках человека обнаружен класс малых ядерных РНК (мяРНК), которые непосредственно не участвуют в синтезе белка, но могут оказывать влияние на процессинг РНК и общую «архитектуру» клетки. Размеры этих относительно небольших молекул варьируют, последние содержат от 90 до 300 нуклеотидов (табл. 37.3).
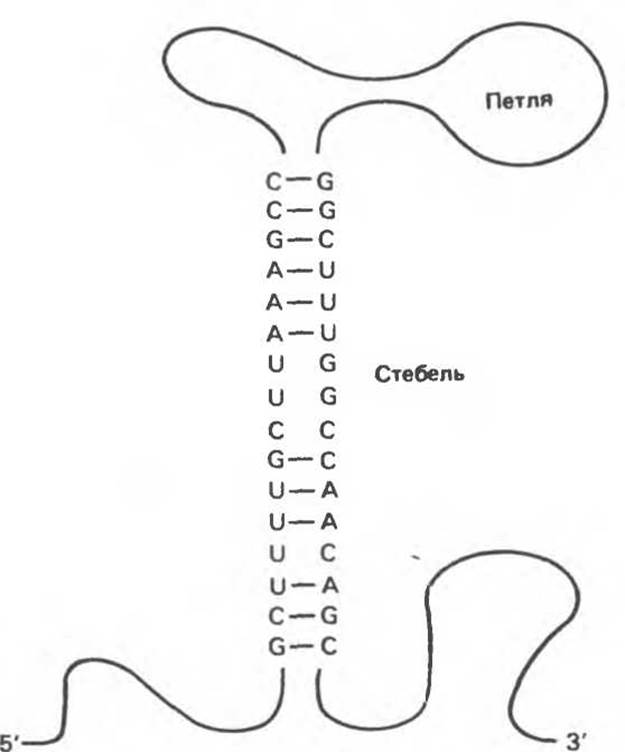
Рис. 37.7. Вторичная структура молекулы РНК типа «петли со стеблем» («шпилька»), возникающая вследствие внутримолекулярного образования водородных связей между комплементарными парами нуклеиновых оснований.
РНК является основным генетическим материалом у некоторых вирусов животных и растений. Некоторые РНК-содержащие вирусы никогда не проходят стадию обратной транскрипции РНК в ДНК. Однако для большинства известных вирусов животных, таких, как ретровирусы, характерна обратная транскрипция их РНК-генома, направляемая РНК- зависимой ДНК-полимеразой (обратной транскриптазой) с образованием двухспиральной ДНК-копии. Во многих случаях образующийся двухспиральный ДНК-транскрипт встраивается в геном и в дальнейшем обеспечивает экспрессию генов вируса, а также наработку новых копий вирусных РНК-геномов.
Структурная организация РНК
Во всех эукариотических и прокариотических организмах существуют три основных класса молекул РНК: информационная (матричная или мессенджер) РНК (мРНК), транспортная (тРНК) и рибосомная (рРНК). Представители этих классов отличаются друг от друга размерами, функциями и стабильностью.
Информационная (мРНК) — наиболее гетерогенный в отношении размеров и стабильности класс. Все представители этого класса служат переносчиками информации от гена к белок-синтезирующей системе клетки. Они выполняют роль матриц для синтезируемого полипептида, т. е. определяют аминокислотную последовательность белка (рис. 37.9).
Информационные РНК, особенно эукариотические, обладают некоторыми уникальными структурными особенностями. 5'-Конец мРНК «кэпирован» 7- метилгуанозинтрифосфатом, присоединенным к 5'-гидроксилу соседнего 2'-О-метилрибонуклеозида через остаток трифосфата (рис. 37.10). Молекулы мРНК часто содержат внутренние остатки 6-метиладенина и 2'-О-метилированные рибонуклеотиды. Хотя смысл «кэпирования» до конца еще не выяснен, можно предположить, что образующаяся структура 5'-конца мРНК используется для специфического узнавания в системе трансляции. Синтез белка начинается на 5'-(кэпированном) конце мРНК. Другой конец большинства молекул мРНК (З'-конец) содержит полиаденилатную цепочку из 20—250 нуклеотидов. Специфические функции этого 3'- ро1у(А)-«хвоста» окончательно не установлены. Можно предполагать, что данная структура отвечает за поддержание внутриклеточной стабильности мРНК. Некоторые мРНК, включая гистоновые мРНК, не содержат poly (А). Наличие poly (А) в структуре мРНК используется для отделения мРНК от других видов РНК посредством фракционирования тотальной РНК на колонках с oligo (Т), иммобилизованным на твердом носителе типа целлюлозы. Связывание мРНК с колонкой происходит за счет комплементарных взаимодействий poly (А)-«хвоста» с иммобилизованным oligo (Т).

Рис. 37.8. Последовательность гена и его РНК-транскрипта. Показаны кодирующая и некодирующая цепи, и отмечена их полярность. РНК-транскрипт, имеющий полярность 5'→3', комплементарен кодирующей цепи (с полярностью 3'→5') и идентичен по последовательности (за исключением замен Т на U) и полярности некодирующей цепи ДНК.
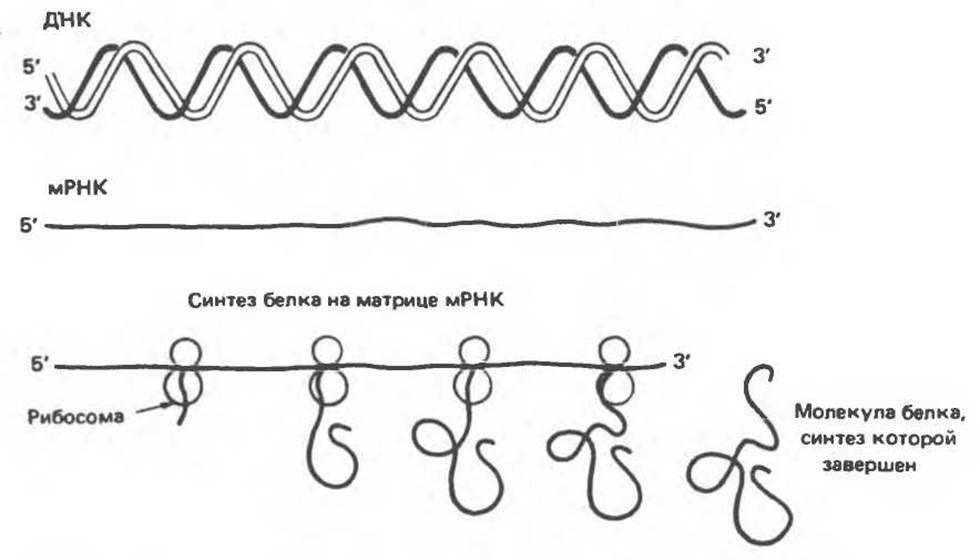
Рис. 37.9. Экспрессия генетической информации ДНК в форме мРНК-транскрипта и последующая трансляция при участии рибосом с образованием специфической молекулы белка.
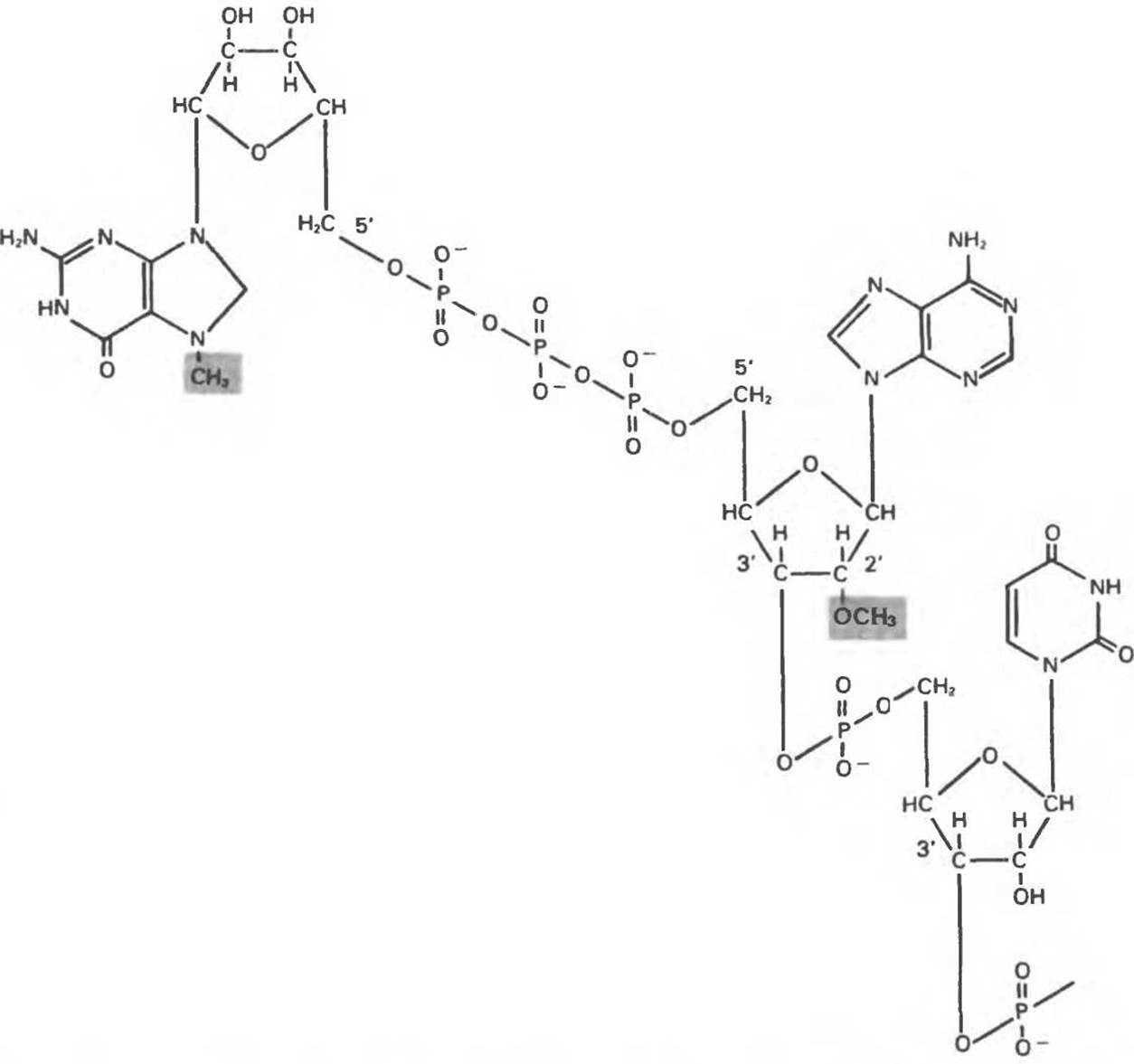
Рис. 37.10. Структура «кэпа», находящегося на 5'-конце большинства эукариотических матричных РНК 7-метилгуанозинтрифосфат присоединяется к 5'-концу мРНК. на котором обычно находится 2'-O-метилпуриновый нуклеотид.
В клетках млекопитающих, включая клетки человека, зрелые молекулы мРНК, находящиеся в цитоплазме, не являются полной копией транскрибируемого участка гена. Образующийся в результате транскрипции полирибонуклеотид представляет собой предшественник цитоплазматической мРНК, перед выходом из ядра он подвергается специфическому процессингу. Непроцессированные продукты транскрипции, обнаруживаемые в ядрах клеток млекопитающих, образуют четвертый класс молекул РНК. Такие ядерные РНК очень гетерогенны и достигают значительных размеров. Молекулы гетерогенных ядерных РНК (гяРНК) могут иметь молекулярную массу более 107, в то время как молекулярная масса мРНК обычно не превышает 2∙106. гяРНК подвергаются процессингу в ядре, и образующиеся зрелые мРНК поступают в цитоплазму, где служат матрицей для биосинтеза белка.
Молекулы транспортных РНК (тРНК) обычно содержат около 75 нуклеотидов. Молекулярная масса таких молекул составляет ~ 25 000. тРНК также формируются в результате специфического процессинга соответствующих молекул-предшественников (см. гл. 39). Транспортные тРНК выполняют функцию посредников в ходе трансляции мРНК. В любой клетке присутствуют не менее 20 видов молекул тРНК. Каждый вид (иногда несколько видов) тРНК соответствует одной из 20 аминокислот, необходимых для синтеза белка. Хотя каждая специфическая тРНК отличается от других нуклеотидной последовательностью, все они имеют и общие черты. Благодаря нескольим внутрицепочечным комплементарным участкам, все тРНК обладают вторичной структурой, получившей название «клеверный лист» (рис. 37.11).
Молекулы всех видов тРНК имеют четыре основных плеча. Акцепторное плечо состоит из «стебля» спаренных нуклеотидов и заканчивается последовательностью ССА (5'→3'). Именно через 3'-гидроксильную группу аденозильного остатка происходит связывание с карбоксильной группой аминокислоты. Остальные плечи тоже состоят из «стеблей», образованных комплементарными парами оснований, и петель из неспаренных оснований (рис. 37.7). Антикодоновое плечо узнает нуклеотидный триплет или кодон (см. гл. 40) в мРНК. D-плечо названо так из-за наличия в нем дигидроуридина, ТψС-плечо названо по последовательности Т-псевдоуридин-С. Дополнительное плечо представляет собой наиболее вариабельную структуру и служит основой классификации тРНК. тРНК класса 1 (75% от общего их числа) обладают дополнительным плечом длиной 3—5 пар оснований. Дополнительное плечо у тРНК-молекул класса 2 состоит из 13—21 пар оснований и часто включает неспаренную петлю.
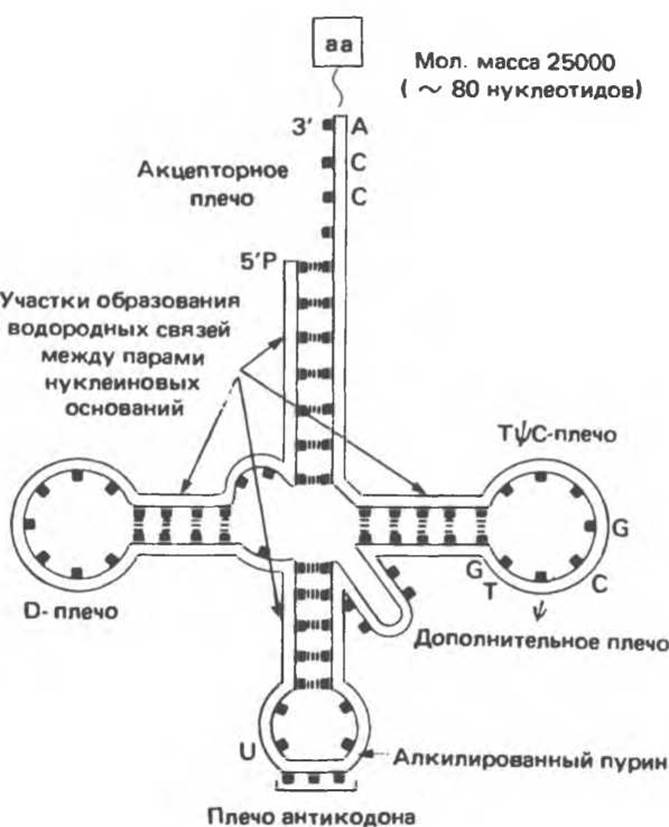
Рис. 37.11. Структура молекулы аминоацил-тРНК, к 3'-ССА-концу которой присоединена аминокислота (аа). Указаны внутримолекулярные водородные связи и расположение анти кодонового, ТТС- и дигидроурацилового (D-) плеч. (From J. D. Watson. Molecular biology of the Gene 3rd, ed., Copyright 1976, 1970, 1965 by W. A. Benjamin, Inc., Menlo Park Calif.)
Вторичная структура, определяемая системой комплементарных взаимодействий нуклеотидных оснований соответствующих плеч, характерна для всех видов тРНК. Акцепторное плечо содержит семь пар оснований, TψC-плечо — пять пар оснований, плечо D — три (или четыре) пары оснований.
Молекулы тРНК весьма стабильны у прокариот и несколько менее стабильны у эукариот. Обратная ситуация характерна для мРНК, которая довольно нестабильна у прокариот, а у эукариотических организмов обладает значительной стабильностью.
Рибосомная РНК. Рибосома — это цитоплазматическая нуклеопротеиновая структура, предназначенная для синтеза белка по мРНК-матрице. Рибосома обеспечивает специфический контакт мРНК и тРНК, в результате которого и происходит трансляция нуклеотидной последовательности, считанной с определенного гена, в аминокислотную последовательность соответствующего белка.
В табл. 37.2 представлены компоненты рибосом млекопитающих, имеющих молекулярную массу 4,2ψ106 и скорость седиментации 80S (единиц Сведберга). Рибосомы млекопитающих состоят из двух нуклеопротеиновых субъединиц — большой с молекулярной массой 2,8∙106 (60S), и малой, имеющей молекулярную массу 1,4∙106 (40S). 60S-субъединица содержит 5S-рибосомную РНК (рРНК), 5,8S-pPHK и 28S-pPHK, а также более 50 различных полипептидов. Малая, 40S-субъединица включает единственную 18S-pPHK и около 30 полипептидных цепей. Все рибосомные РНК, за исключением 5S-PHK, имеют общего предшественника — 45S-PHK, локализованную в ядрышке (см. гл. 40). У молекулы 5S-PHK предшественник собственный. В ядрышке происходит упаковка высокометилированных рибосомных РНК с рибосомными белками. В цитоплазме рибосомы достаточно устойчивы и способны осуществлять большое число циклов трансляции.
Таблица 37.2. Компоненты рибосом млекопитающих 1)
|
Компонент |
Молекулярная масса |
Белковые компоненты |
РНК-компоненты |
|||
|
Число |
Молекулярная масса |
Размер |
Молекулярная масса |
Число оснований |
||
|
40S-субъединица |
1,4∙106 |
~35 |
7∙105 |
18S |
7∙105 |
1900 |
|
60S-субъединица |
2,8∙106 |
~ 50 |
1∙106 |
5S |
35000 |
120 |
|
5,8S |
45000 |
160 |
||||
|
28S |
1,6∙106 |
4700 |
||||
Субъединицы рибосом классифицируют по скорости седиментации в единицах Сведберга (40S и 60S); в таблице указаны масса обеих субъединиц, число индивидуальных белков и их масса, для РНК-компонентов каждой субъединицы приведены размер (единицы Сведберга), молекулярная масса и число оснований
Небольшие стабильные РНК. В эукариотических клетках обнаружено большое число дискретных, высококонсервативных, небольших и стабильных молекул РНК. Большинство РНК этого типа обнаруживаются в составе рибонуклеопротеинов и локализованы в ядре, цитоплазме или одновременно в обоих компартментах. Размеры этих молекул варьируют от 90 до 300 нуклеотидов, содержание их — 100000—1000000 копий на клетку.
Малые ядерные нуклеопротеиновые частицы (часто называемые snurps — от англ. small nuclear ribonucleic particles), вероятно, играют существенную роль в регуляции экспрессии генов. Нуклеопротеиновые частицы типа U7, по-видимому, участвуют в формировании 3'-концов гистоновых мРНК. Частицы U4 и U6, вероятно, необходимы для полиаденилирования, a U1 — для удаления интронов и процессинга мРНК (см. гл. 39). Табл. 37.3. суммирует некоторые характеристики небольших стабильных РНК.
Таблица 37.3. Некоторые виды небольших стабильных РНК, обнаруженные в клетках млекопитающих
|
Наименование |
Длина (число нуклеотидов) |
Число молекул на клетку |
Локализация |
|
U1 |
165 |
1∙106 |
Нуклеоплазма (гяРНК) |
|
U2 |
188 |
5∙105 |
Нуклеоплазма |
|
U3 |
216 |
3∙105 |
Ядрышко |
|
U4 |
139 |
1∙105 |
Нуклеоплазма |
|
U5 |
118 |
2∙105 |
Нуклеоплазма |
|
U6 |
106 |
3∙105 |
Перихроматиновые гранулы |
|
4,5S |
91—95 |
3∙105 |
Ядро и цитоплазма |
|
7S |
280 |
5∙105 |
Ядро и цитоплазма |
|
7-2 |
290 |
1∙105 |
Ядро и цитоплазма |
|
7-3 |
300 |
2∙105 |
Ядро |
Литература
Darnell J. et al. Molecular Cell Biology, Scientific American Books, 1986.
Hunt T. DNA Makes RNA Makes Protein, Elsevier, 1983. Lewin В. Genes, 2nd ed., Wiley, 1985.
Rich A. et al. The chemistry and biology of left-handed Z-DNA, Annu. Rev. Biochem., 1984, 53, 847.
Turner P. Controlling roles for snurps, Nature, 1985, 316, 105. Watson J. D. The Double Helix, Atheneum, 1968.
Watson J. D., Crick F.H.C. Molecular structure of nucleic acids. Nature, 1953, 171, 737.
Zieve G. W. Two groups of small stable RNAs, Cell, 1981, 25, 296.