Биохимия человека Том 2 - Марри Р. 1993
Структура, функция и репликация информационных макромолекул
Нуклеотиды
Природные нуклеотиды
Свободные нуклеотиды также выполняют важные функции в различных тканях организма (см. ниже).
Производные аденозина
ADP и АТР являются субстратом и продуктом окислительного фосфорилирования (гл. 3). АТР выполняет функцию основного внутриклеточного переносчика свободной энергии (гл. 11). Концентрация наиболее распространенного свободного нуклеотида в клетках млекопитающих — АТР—составляет около 1 ммоль/л.
Циклический АМР (3', 5'-аденозинмонофосфат, сАМР) — медиатор различных внеклеточных сигналов в клетках животных — образуется из АТР в результате реакции, катализируемой аденилатциклазой (рис. 34.14). Активность аденилатциклазы регулируется комплексом взаимодействий, многие из которых инициируются через рецепторы гормонов (гл. 43). Внутриклеточная концентрация сАМР (около 1 мкмоль/л) на 3 порядка ниже концентрации АТР. Гидролиз сАМР до 5'-АМР катализирует сАМР-фосфодиэстераза (рис. 34.14).
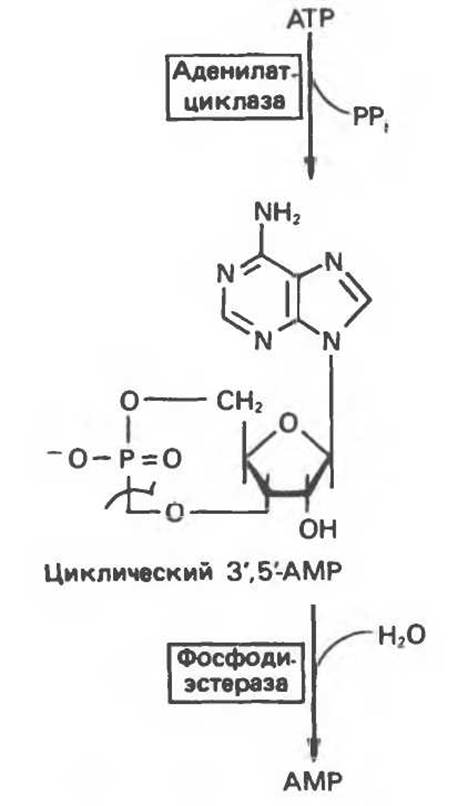
Рис. 34.14. Образование сАМР из АТР и гидролиз сАМР фосфодиэстеразой.
Включение остатка сульфата при образовании таких соединений, как сульфатированные протеогликаны (гл. 42), требует его предварительной активации (рис. 34.15). Сульфат активируется в реакции с АТР, образуя аденозин-3'-фосфат-5'-фосфосульфат. Активированный сульфат необходим также как субстрат реакции образования сульфатных конъюгатов.
S-Аденозилметионин (рис. 34.16) представляет собой «активную» форму метионина. S-аденозилметионин выполняет функцию донора метильной группы во многих реакциях метилирования и, кроме того, является источником пропиламина для синтеза полиаминов.
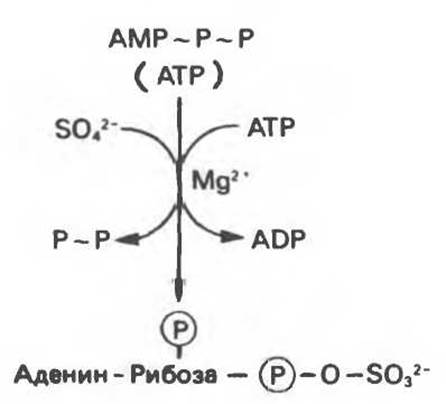
Рис. 34.15. Образование аденозин-3'-фосфат-5'-фосфосульфата.
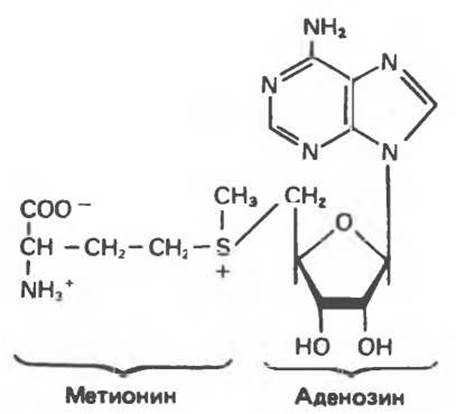
Рис. 34.16. S-Аденозилметионин.
Производные гуанозина
Нуклеотиды этого типа, в частности гуанозиндифосфат и гуанозинтрифосфат, участвуют в нескольких, требующих энергии, биохимических процессах, где они выступают функциональными аналогами ADP и АТР. Например, окисление а-кетоглутаровой кислоты до сукцинил-СоА в цикле трикарбоновых кислот сопровождается фосфорилированием GDP до GTP. GTP необходим для активации аденилатциклазы некоторыми гормонами; он выполняет функции как аллостерического регулятора, так и источника энергии в процессе синтеза белка на полирибосомах. Таким образом, GTP играет важную роль в поддержании внутриклеточного энергетического баланса.
Циклический GMP (3’, 5’-гуанозинмонофосфат, cGMP) (рис. 34.17) служит внутриклеточным проводником внеклеточных сигналов. В некоторых случаях cGMP выступает в роли антагониста с АМР. cGMP образуется из GTP под действием гуанилатциклазы — фермента, имеющего много общего с аденилатциклазой. Гуанилатциклаза, как и аденилатциклаза, регулируется различными эффекторами, в том числе и гормонами. Как и cAMP, cGMP гидролизуется фосфодиэстеразой до соответствующего 5’-монофосфата.
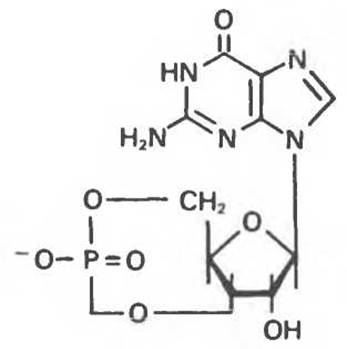
Рис. 34.17. Циклический 3'5'-гуанозинмонофосфат (cGMP).
Производные гипоксантина
Гипоксантиновый рибонуклеотид, обычно называемый инозиновой кислотой (IMP или инозинат в солевой форме), — представляет собой предшественник всех пуриновых рибонуклеотидов, синтезируемых de novo. Инозинат может образовываться в реакции дезаминирования АМР. Эта реакция происходит главным образом в мышечной ткани и является частью цикла пуриновых нуклеотидов. Функционирование этой части цикла приводит в конечном итоге к образованию аммиака за счет аспартата (рис. 34.18). При удалении фосфатной группы из IMP образуется нуклеозидное производное инозина (гипоксантинрибозид) — промежуточное соединение цикла «реутилизации» пуринов (гл. 35).
Инозиндифосфат (IDP) и инозинтрифосфат (ITP) представляют собой аналоги ADP и АТР, у которых в качестве пуринового нуклеозида выступает инозин; они иногда принимают участие в реакциях фосфорилирования.
Производные урацила
Производные урациловых нуклеотидов участвуют в качестве коферментов в реакциях метаболизма гексоз и полимеризации углеводов, в частности при биосинтезе крахмала и олигосахаридных фрагментов гликопротеинов и протеогликанов (гл. 54). Субстратами в этих реакциях являются уридиндифосфатсахара. Например, уридиндифосфатглюкоза (UDPGlc) служит предшественником гликогена. Другой кофермент — уридиндифосфатглюкуроновая кислота (UDPGlcUA) — выполняет функцию «активного» глюкуронида в реакциях конъюгирования, например при образовании глюкуронида билирубина (гл. 33).
Урацил участвует в образовании макроэргических фосфатных соединений, аналогичных АТР, GTP, ІТР. Уридинтрифосфат (UTP) используется, например, в реакции превращения галактозы в глюкозу; при этом образуются UDPGlc и UDPGal. Кроме того, UTP служит одним из мономерных предшественников РНК.
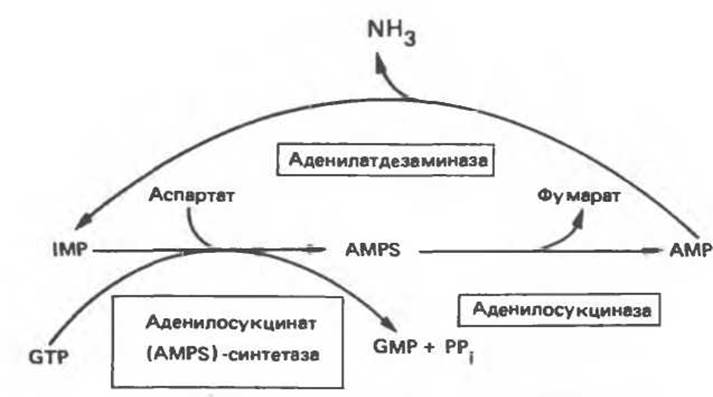
Рис. 34.18. Цикл пуриновых нуклеотидов.
Производные цитозина
Цитидин (цитозинрибозид) может формировать высокоэнергетические фосфатные соединения — цитидиндифосфат (CDP) и цитидинтрифосфат (СТР). Последний выступает также в роли предшественника при включении СМР в состав нуклеиновых кислот. СТР необходим для биосинтеза некоторых фосфоглицеридов в тканях животных. Реакции с участием церамида и CDP-холина приводят к образованию сфингомиелина и других замещенных сфингозинов. Известны циклические производные цитидина, аналогичные вышеописанным циклическим производным аденозина и гуанозина.
Нуклеотиды в составе коферментов
Функциональными фрагментами многих коферментов являются нуклеотиды, структурно аналогичные пуриновым и пиримидиновым нуклеотидам (табл. 34.1).
Таблица 34.1. Многие коферменты и родственные им соединения являются производными аденозинмонофосфата
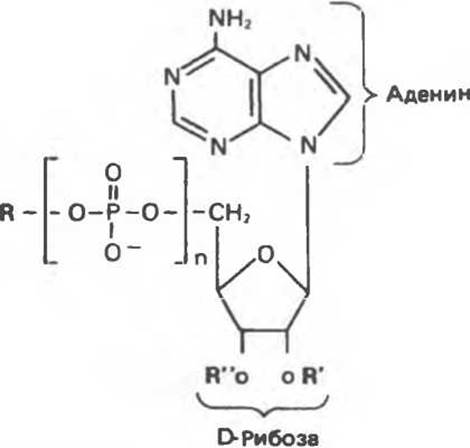
|
Кофермент |
R |
R' |
R" |
n |
|
Активный метионин |
Метионин 1) |
Н |
Н |
0 |
|
Аденилаты аминокислот |
Аминокислота |
Н |
Н |
1 |
|
Активный сульфат |
SO2 |
Н |
РО2-3 |
1 |
|
3'-5'-сАМР |
Н |
Н |
РО2-3 |
1 |
|
NAD |
* |
Н |
Н |
2 |
|
NADP |
* |
РО2-3 |
Н |
2 |
|
FAD |
* |
Н |
Н |
2 |
|
СоА∙SH * |
Н |
РО2-3 |
2 |
|
1) Замещает фосфатную группу
* R —производное витамина В