Биохимия аминокислот - А. Майстер 1961
Роль аминокислот в питании
Потребность в аминокислотах у высших животных
Более 40 лет назад Осборн и Мендель [1] установили, что триптофан и лизин являются пищевыми факторами, необходимыми для роста крыс. Эти классические исследования, в которых были использованы рационы из очищенных белков, послужили толчком для дальнейших работ, показавших насущную роль гистидина [2] и цистина [3]. После открытия в 1935 г. треонина [4] появилась возможность применять рационы, содержащие вместо белков смеси аминокислот. Роуз и его сотрудники [5] установили путем последовательного исключения аминокислот, из рациона поодиночке, что для роста молодых крыс незаменимыми являются 10 аминокислот. В табл. 10 приведены данные о потребности в аминокислотах у 9 видов животных. Эти данные интересны во многих отношениях. Аланин, аспарагиновая кислота, оксипролин и серин не являются незаменимыми ни для одного из видов высших животных, из чего можно заключить, что они синтезируются в организме. Аргинин не необходим для сохранения азотистого равновесия у взрослых людей, собаки, крысы и мыши. У молодых крыс аргинин необходим для оптимального роста, хотя некоторый рост наблюдается и на рационах, лишенных этой аминокислоты [19]. В противоположность крысам цыплята не могут синтезировать аргинин [20]. Аргинин удается заменить в питании молодых цыплят цитруллином, но не орнитином [21]. Для обеспечения оптимального роста цыплят необходимы также глицин, глутаминовая кислота и пролин.
Гистидин необходим для роста крысы, собаки, мыши и цыпленка. Он нужен, по-видимому, и для сохранения азотистого равновесия у взрослых крыс [22, 23], хотя более ранние исследования указывали на его заменимость [24]. Для сохранения азотистого равновесия у человека гистидин не нужен [12].
Восемь аминокислот (лизин, треонин, триптофан, метионин, фенилаланин, лейцин, валин и изолейцин) нербходимы для всех исследованных видов животных (см. табл. 10). Исследования в области питания во многом способствовали выяснению путей обмена этих аминокислот. Ряд примеров будет приведен здесь и в гл. IV.
Фенилаланин незаменим для крыс и цыплят, но необходимое количество его в рационе может быть значительно снижено за счет добавления к рациону тирозина [25, 26]. Сберегающее действие тирозина связано с установленным в ряде других исследований превращением фенилаланина в тирозин (стр. 417). Обратная реакция, т. е. образование фенилаланина из тирозина, у высших животных, по-видимому, не происходит [27], вследствие чего тирозин не может полностью заместить фенилаланин в питании.
Потребность крыс в метионине может быть покрыта за счет приема с пищей гомоцистина при условии введения в рацион донаторов «лабильных» метальных групп или достаточного количества фолевой кислоты и витамина B12 [28, 29]. Цистин оказывает «сберегающее» влияние на потребность в метионине [30—32]. L-Цистатионин может заменять в питании цистин, a L-алло-цистатионин, превращающийся в организме в гомоцистеин, поддерживает рост в тех же условиях, что и гомоцистеин [33]. Эти факты вполне понятны в свете имеющихся данных об обмене упомянутых аминокислот (стр. 367).
Таблица 10 Потребность в аминокислотах у животных различных видов*
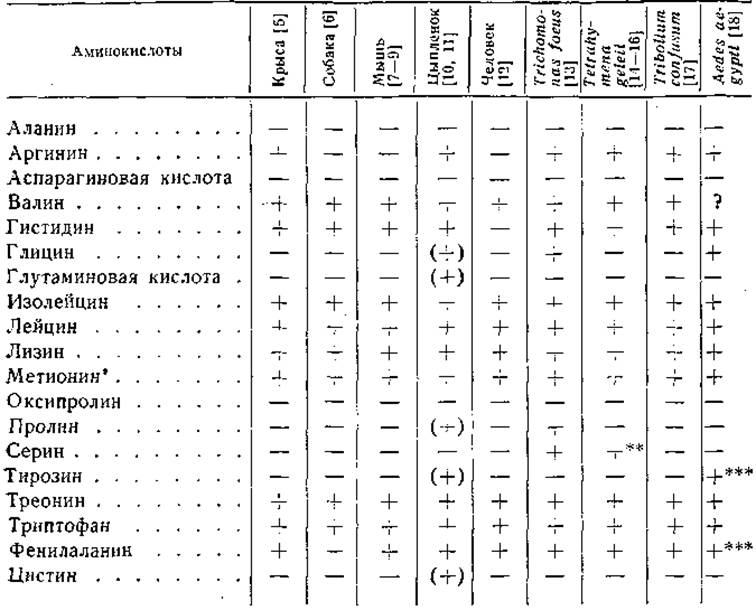
* + незаменима; — заменима; (+) необходима в определенных условиях. У всех животных, кроме человека, критерием служит рост; у человека критерием служит обеспечение азотистого равновесия.
** Потребность в серине зависит от штамма.
*** Необходимо одновременное наличие тирозина и фенилаланина (см. стр. 133).
При помощи опытов по кормлению животных были выяснены взаимосвязи аргинина, пролина и глутаминовой кислоты в обмене веществ. Глутаминовая кислота или пролин могут частично замещать аргинин как фактор, повышающий скорость роста молодых крыс [32].
В исследованиях по аминокислотному питанию обычно пренебрегали значением кишечной флоры. Бактерии кишечника играют заметную роль в синтезе витаминов, однако не доказано, что они выполняют такую же функцию в отношении аминокислот. Исследования, проведенные с применением антибактериальных средств, не показали сколько-нибудь заметного влияния этих соединений [34—36]. Четкий ответ на этот вопрос можно было бы получить в опытах, проводимых в условиях полной стерильности животных (ср. [29]). Использование аминокислот при их парентеральном введении установлено как у лабораторных животных, так и у человека. Возможно, что бактерии кишечника в какой-то мере разрушают принятые с пищей аминокислоты и другие азотистые соединения (см., например, данные о роли бактериальной уреазы, стр. 173), однако до сих пор не получено данных о сколько-нибудь значительной роли синтеза аминокислот бактериями в питании животных. Полагают, что образование значительных количеств аминокислот при самопереваривании протеолитических ферментов в желудочно-кишечном тракте может играть некоторую роль, обеспечивая наличие в кишечнике смеси аминокислот относительно постоянного состава [290]. Такую же роль, очевидно, может играть действие пищеварительных ферментов на бактериальную флору.
Роуз и его сотрудники [12,37—52] исследовали количественно потребность в аминокислотах для сохранения азотистого равновесия у здоровых взрослых мужчин. Испытуемые получали рацион, состоявший из чистых аминокислот, маисового крахмала, сахара, коровьего и кукурузного масла, рыбьего жира, неорганических солей, центрифугированного лимонного сока, муки и витаминов. В этом рационе на аминокислоты приходилось 95%. общего количества азота, главным источником остального азота служили примеси в препарате крахмала. После установления азотистого равновесия ту или другую аминокислоту исключали из рациона, причем постоянство общего количества вводимого с пищей азота обеспечивалось увеличением количества других аминокислот. Таким путем была установлена незаменимость восьми аминокислот (см. табл. 10). Значительный интерес представляет установление того факта, что гистидин не является необходимым для обеспечения азотистого равновесия у человека. Отсюда можно заключить, что обмен гистидина у крысы (и у других животных) существенным образом отличается от его обмена у человека. Олбениз и сотрудники [53] нашли, что азотистое равновесие у людей может быть сохранено без введения гистидина, однако испытуемые в этих условиях теряют в весе; Роуз и сотрудники [42] не наблюдали у испытуемых потери веса при условии введения с пищей большего количества калорий. Следует также иметь в виду, что гистидин может образоваться в известных условиях при распаде гемоглобина [55]. Известно, что гистидин играет большую роль в регенерации гемоглобина у крыс, подвергнутых кровопусканию [54]. В кратковременных опытах на крысах, не получавших с пищей гистидина, сохранению азотистого равновесия сопутствовал распад гемоглобина [55].
Используя описанный выше метод, Роуз и сотрудники [43—52] определили количества аминокислот, необходимые для сохранения азотистого равновесия у молодых взрослых мужчин. В состав рациона в качестве добавочных источников азота включали глицин, мочевину или оба этих соединения; с целью повысить калорийность рациона до 55 кал на 1 кг веса тела добавляли сливочное масло и сахар. Этим способом были получены данные, приведенные в табл. 11. Число обследованных людей было сравнительно невелико (все они были примерно одного возраста), и впредь до проведения более широких исследований полученные данные нужно рассматривать как предварительные. Следует также отметить, что опытный рацион давали в течение сравнительно короткого срока и что единственным критерием при этом служило сохранение азотистого равновесия.
Таблица 11 Потребность в аминокислотах у человека [52] *
|
Аминокислота |
Число количественных опытов |
Экспериментально установленные пределы потребности, г в день |
Количество, условно принятое за минимальное, г в день |
«Безусловно достаточное количество, г в день |
Число людей, сохранявших азотистое равновесие при приеме «достаточных« или меньших количеств аминокислот |
|
L-Триптофан |
3 |
0,15—0,25 |
0,25 |
0,50 |
42 |
|
L-Фенилаланин |
6 |
0,80—1,10 |
1,10 |
2,20 |
32 |
|
L-Лизин |
6 |
0,40—0,80 |
0,80 |
1,60 |
37 |
|
L-Треонин |
3 |
0,30—0,50 |
0,50 |
1,00 |
29 |
|
L-Валин |
5 |
0,40—0,80 |
0,80 |
1,60 |
33 |
|
L-Метионин |
6 |
0,80—1,10 |
1,10 |
2,20 |
23 |
|
L-Лейцин |
5 |
0,50—1,10 |
1,10 |
2,20 |
18 |
|
L-Изолейцин |
4 |
0,65—0,70 |
0,70 |
1,40 |
17 |
* Все величины определены при использовании рационов, содержащих 8 незаменимых аминокислот и добазочный азот в количестве, достаточном для синтеза заменимых аминокислот.
Роуз и сотрудники нашли, что установленная ими минимальная потребность в L-метионине может быть покрыта на 80—89% введением в рацион L-цистина и что 70—75% потребного L-фенилаланина удается заменить L-тирозином. Сберегающее влияние тирозина и цистина на потребность в фенилаланине и соответственно метионине было несколько большим, чем в аналогичных опытах на крысах. Наконец, Роуз и сотрудники [51] в своих исследованиях установили, что количество вводимого с пищей азота, необходимое для синтеза заменимых аминокислот, не превышает 2,55 г в день.
Потребности в аминокислотах у младенцев были изучены Олбенизом [20]. О потребности в определенной аминокислоте судили по тому, какое количество ее необходимо для обеспечения нормального прироста веса и усвоения азота у ребенка, получавшего ранее недостаточное питание. Эти. исследования показали, что гистидин и аргинин, по-видимому, не существенны для питания младенцев мужского пола, что незаменимыми являются те восемь аминокислот, которые незаменимы в питании взрослых людей, и что в известных условиях проявляется потребность в цистине и тирозине. Данные о потребности в аминокислотах у младенцев и у взрослых сопоставлены в табл. 12. Хотя эти данные носят предварительный характер и требуют дальнейшего подтверждения, интересно отметить, что младенцы нуждаются в относительно больших количествах лизина, треонина и валина, чем взрослые. Весьма любопытно, что относительная потребность в изолейцине у взрослых и у младенцев почти одинакова, тогда как потребность в лейцине у последних значительно выше.
Есть основание думать, что потребности организма в аминокислотах должны в количественном отношении соответствовать аминокислотному составу синтезируемых белков. Так, например, отмечено близкое соответствие между потребностями в аминокислотах у крыс и цыплят и соотношением аминокислот в основных тканях тела этих животных [56—61]. Эти данные говорят о том, что совокупность механизмов синтеза белка, которыми располагает организм, обеспечивает эффективное использование имеющихся аминокислот. Изменения содержания аминокислот в пище не отражаются на аминокислотном составе белков тела крысы и цыпленка. Если аминокислоты вводятся с пищей в количестве, близком тому, которое необходимо для обеспечения синтеза белка, то содержание аминокислот в крови возрастает лишь незначительно; введение аминокислот в избытке сопровождается повышением их содержания в крови [62].
Таблица 12 Сопоставление потребностей взрослых мужчин [43—52] и младенцев [20] в отношении незаменимых аминокислот пищи
|
Аминокислота |
Взрослые мужчины |
Младенцы мужского пола |
||
|
потребность, мг/кг |
относительное значение для азотистого равновесия * |
потребность, мг/кг |
относительное значение для роста * |
|
|
Триптофан |
7,2 |
1,0 |
30 |
1,0 |
|
Фенилаланин |
31 |
4,3 |
169 |
5,6 |
|
Лизин |
23 |
3,2 |
170 |
5,6 |
|
Треонин |
14 |
1,9 |
87 |
2,9 |
|
Валин |
23 |
3,2 |
161 |
5,4 |
|
Метионин |
31 |
4,3 |
85 |
2,8 |
|
Лейцин |
31 |
4,3 |
425 |
14,0 |
|
Изолейцин |
20 |
2,8 |
90 |
3,0 |
* При расчете относительного значения потребности в каждой аминокислоте для сохранения азотистого равновесия и обеспечения роста за единицу приняты величины потребности в триптофане»
Потребность в аминокислотах возрастает во время беременности и лактации, а также при некоторых заболеваниях и после травмы, так как при этом аминокислоты расходуются для добавочного синтеза клеток крови, белков молока, антител и т. д. (ср. [63]). Иногда возникает потребность в больших, чем обычно, количествах определенных аминокислот. Например, для обильного роста шерсти у овец необходимо введение с кормом относительно больших количеств серусодержащих аминокислот. При фенилпировиноградной олигофрении к рациону с низким содержанием ароматических аминокислот необходимо добавлять тирозин, в котором здоровые люди не нуждаются; поэтому тирозин следует считать незаменимой аминокислотой при этом заболевании (стр. 474).