Биохимия - Химические реакции в живой клетке Том 1 - Д. Мецлер 1980
Как молекулы соединяются друг с другом
Упаковка макромолекул
Олигомеры, обладающие циклической симметрией
Аналогично тому как аминокислоты, сахара и нуклеотиды служат «строительными блоками» для белков, полисахаридов и нуклеиновых кислот, так и сами эти макромолекулы в свою очередь являются единицами, из которых собираются более сложные структуры. Волокна, микротрубочки, оболочки вирусов и небольшие симметричные группы субъединиц в олигомерных ферментах — все это варианты строго упорядоченной упаковки макромолекул (которую иногда называют четвертичной структурой). Рассмотрим сначала наиболее простой случай агрегации идентичных белковых субъединиц. Известно, что, хотя форма многих белков близка к сферической, тем не менее они не совсем симметричны. На приведенных ниже рисунках это их свойство несколько преувеличено, чтобы более четко проиллюстрировать общие принципы упаковки.
На рис. 4-6 схематически изображены субъединицы (протомеры), содержащие взаимно комплементарные участки а и j. Две такие молекулы стремятся соединиться друг с другом так, чтобы участок а одного протомера оказался связанным с участком j другого. При этом с одной стороны димера остается свободный участок а, a с другой — свободный участок j, к которым могут присоединяться другие протомеры. В некоторых случаях таким способом образуются длинные цепи.
Если геометрия молекул соответствует схеме, показанной на рис. 4-6, А, то к димеру может легко «пристроиться» третья субъединица и замкнуть кольцо с образованием тримера. В зависимости от геометрии субъединиц кольцо может быть меньшим (димер) или большим (тетрамер, пентамер и т. д.). Связь, в которой участвуют различные участки (а и j) субъединицы, иногда называют гетерологической [33]. Для образования замкнутого кольца, состоящего из нескольких субъединиц, необходимо, чтобы между связывающимися участками (а, j) был вполне определенный угол.
Кольцо, образованное за счет только одних гетерологических взаимодействий, должно обладать циклической симметрией. Так, например, показанный на рис. 4-6, А тример имеет ось симметрии третьего порядка. Это означает, что каждая субъединица может быть совмещена со следующей поворотом вокруг оси симметрии на 360°/3. Симметрия такого олигомера обозначается символом С3. Субъединичная структура многих природных белков, включая все белки, состоящие из трех, пяти или любого другого нечетного числа одинаковых протомеров, обладает, по-видимому, циклической симметрией.
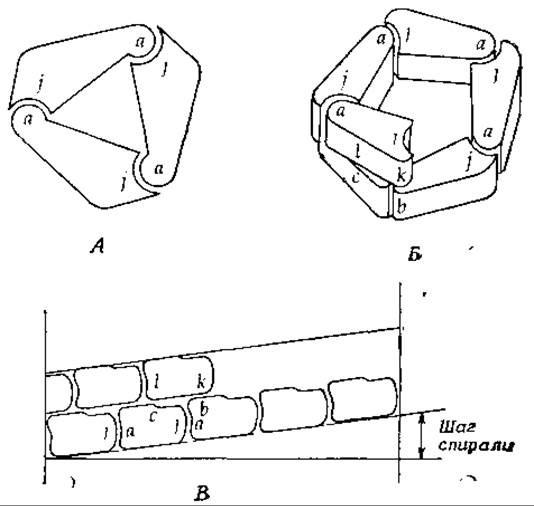
РИС. 4-6. Гетерологическое связывание субъединиц. А — кольцо; Б — спираль; В — «радиальная проекция» субъединиц, образующих спираль Б. Участки связывания субъединиц обозначены буквами а, b, с, j, k и I.
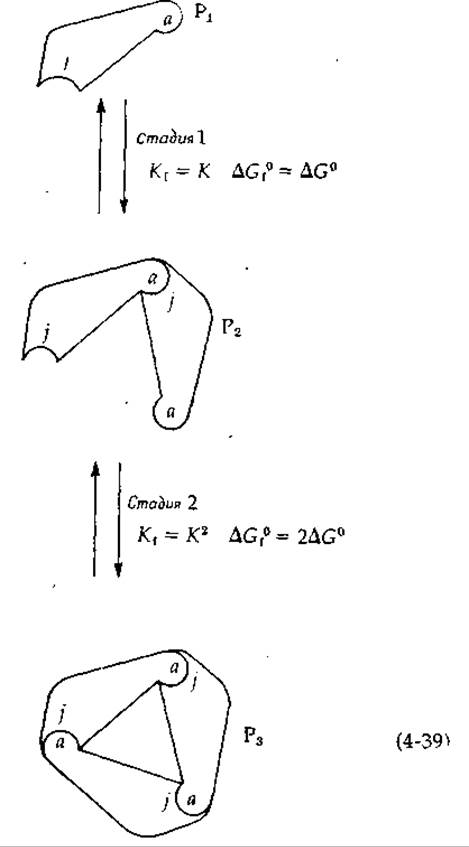
Рассмотрим теперь количественные аспекты гетерологических взаимодействий, приводящих к образованию замкнутых колец. Пусть К — константа образования, a ∆G0 — изменение стандартной свободной энергии для присоединения j-конца протомера Р к a-концу другого протомера Р, в результате чего образуется димер Р2 [уравнение (4-39)]. На второй стадии присоединяется третий протомер. Заметим, что при этом возникают два новых аj-контакта, в связи с чем ∆G0f для второй стадии равно 2∆G0, a K1— К2. Результирующая константа образования для процесса сборки тримера из трех протомеров и изменение свободной энергии равны1

1 Предполагается, что значения ∆G° образования обеих новых аj-связей в тримере одинаковы и равны ∆G0образования aj-связи в димере. Читатель вправе усомниться в правильности такого предположения и подумать о том, при каких условиях значение Kf, вычисленное для тримера в соответствии с уравнением (4-40), окажется завышенным или заниженным.
Рассмотрим теперь гипотетический случай, когда протомер Р непрерывно синтезируется в клетке и сразу же распадается в результате параллельно протекающей метаболической реакции на неспособные к ассоциации продукты. При этом обе реакции сбалансированы таким образом, что стационарная концентрация протомера [Р] остается все время равной 10-5 М. Предположим, что значение К для единичного aj-взаимодействия, приводящего к образованию димеров и тримеров (колец), равно 104 (или ∆G0 = —22,8 кДж∙моль-1). Спрашивается: чему будет равна концентрация димеров и тримерных колец в клетке в равновесии, если концентрация Р равна 10-5 М? Используя уравнение (4-39), получаем, что концентрация димеров Р2 равна 104∙(10-5)2 = 10-6 М. (Заметим, что при такой концентрации димеров концентрация составляющих их мономеров эквивалентна 2∙10-6 М.) Концентрация колец, [Р3], равна (104)3∙(10-5)3 = 10-3 М (концентрация соответствующих мономерных единиц эквивалентна 3∙10-3 М), Таким образом, 99,6% суммарного количества протомеров Р, присутствующих в клетке, будет входить в состав тримеров (10-5+0,2∙10-5+300∙10-5 М), 0,33% — в состав мономеров и только 0,07% — в состав димеров. Следовательно, одновременное образование двух гетерологических связей, приводящее к замыканию кольца, делает реакцию ассоциации, описываемую уравнением (4-39), высококооперативным процессом. В клетке мы обнаружим в основном кольца и мономеры, но не димеры.
Как изменится относительное содержание небольших колец внутри клетки, если скорость превращения протомера Р в форму, не способную к ассоциации, вдруг сильно возрастет, так что концентрация Р упадет до 10-6 М? Определите сами равновесные концентрации мономерных единиц Р1 димеров Р2 и тримеров Р3, если К останется при этом равной 104. Здесь и проявится особенность кооперативных процессов, которая состоит в том, что они характеризуются зависимостью от концентрации не в первой, а в более высокой степени.