Биохимия - Химические реакции в живой клетке Том 1 - Д. Мецлер 1980
Мембраны и клеточные оболочки
Стенки растительных клеток
Толстая стенка растительной клетки (рис. 1-3) устроена необычайно сложно [113—116]. Благодаря ее сложному строению растения обладают прочностью и жесткостью, а их клетки способны к быстрому удлинению в период роста. Норткот [113] сравнил строение стенки растений с фибраглассом — пластиком, армированным стекловолокном. Так, в стенке клетки находятся микрофибриллы, состоящие из целлюлозы и других полисахаридов, которые погружены в матрикс, также состоящий в основном из полисахаридов. На ранних стадиях роста зеленых растений закладывается первичная клеточная стенка, содержащая свободно переплетенные целлюлозные волокна диаметром приблизительно 10 нм, центральная часть которых (~4 нм) имеет кристаллическую структуру. Такие целлюлозные волокна содержат 8000—12 000 остатков глюкозы.
В процессе роста растения под первичной стенкой образуется вторичная клеточная стенка, которая построена из многочисленных слоев плотно упакованных микрофибрилл, имеющих различное направление в соседних слоях. Микрофибриллы зеленых растений чаще всего состоят из целлюлозы, однако могут содержать также и другие полисахариды. Фибриллы некоторых водорослей богаты ксиланом и маннаном.
В период роста растения состав матрикса меняется. На начальных стадиях развития матрикс в основном состоит из пектиновых веществ (производных полигалактуроновой кислоты), однако в дальнейшем в нем появляются ксиланы и различные нейтральные полисахариды («гемицеллюлоза»). В состав первичной клеточной стенки двудольных растений входят ксилоглюканы (содержащие линейную глюкановую цепь и боковые цепи из остатков ксилозы, галактозы и фукозы), арабиногалактаны и рамногалактоуронаны (состоящие из прямых цепей, построенных из остатков галактуроновой кислоты, между которыми через определенные интервалы включены рамнозные кольца). Эти три сложных полисахарида ковалентно связаны друг с другом и с целлюлозными микрофибриллами [114] (рис. 5-12).
Главными компонентами клеточных стенок растений являются углеводы. Однако кроме углеводов в них содержатся небольшие количества гликопротеида, получившего название экстензии [117]. Это соединение, подобно коллагену внутриклеточного матрикса тканей животных, содержит в большом количестве 4-оксипролин. В него входят также арабиноза и галактоза, присоединенные в форме олигосахаридов (состоящих приблизительно из 9 остатков) к гидроксильным группам белка. На более поздних стадиях развития, когда стенки затвердевают до состояния древесины, в клетках растений откладываются значительные количества лигнинов (гл. 14, разд. 3.6). Эти химически устойчивые полимеры содержат большое число ароматических колец.
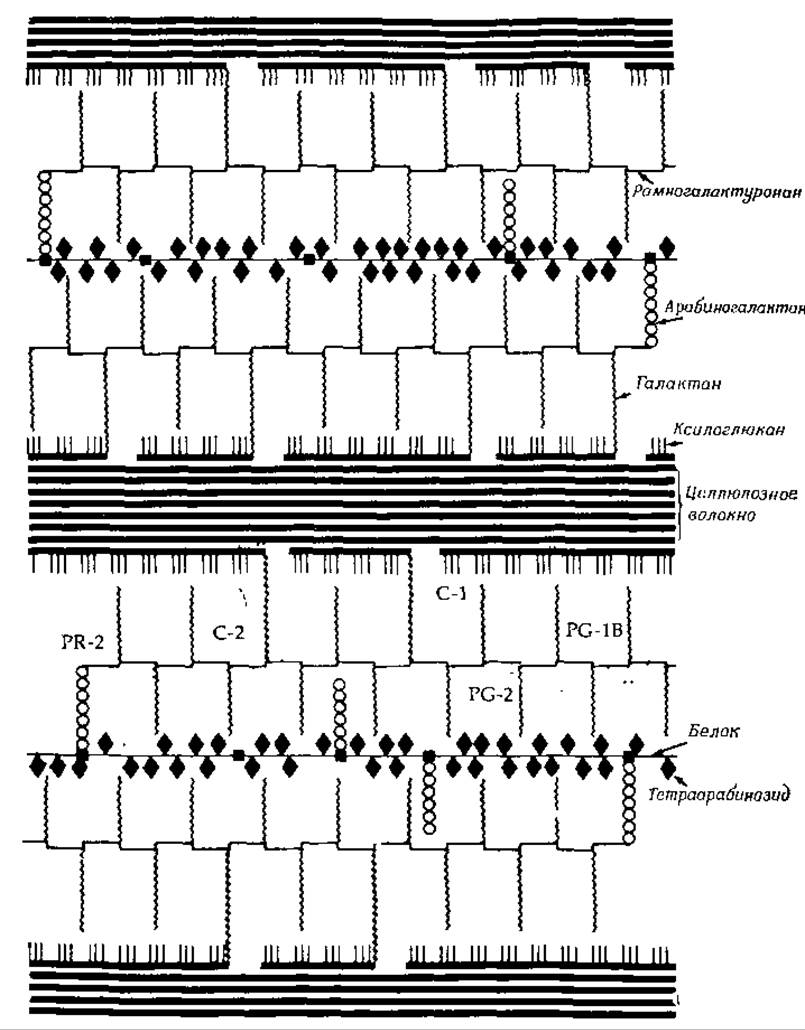
РИС. 5-12. Предполагаемое строение стенки растительных клеток. Относительные размеры различных компонентов стенки не соблюдены, однако соотношения между ними близки к истинным. Расстояние между пучками целлюлозных волокон увеличено, чтобы можно было изобразить соединяющие структуры В первичной клеточной стенке одной клетки может находиться от 10 до 100 пучков целлюлозных волокон [115]
Биосинтез пектинов и гемицеллюлоз происходит, вероятно, в пузырьках аппарата Гольджи, откуда эти соединения транспортируются наружу путем экзодитоза. Что же касается целлюлозных волокон, то они могут выталкиваться через плазматическую мембрану. Интересным свойством первичных клеточных стенок растений является их способность чрезвычайно быстро удлиняться во время роста клетки. Увеличение размера клеточных стенок, так же как и общее увеличение объема клетки, обусловлено ростом давления внутри клетки. Характер увеличения строго регулируется после определенного этапа развития; под влиянием ряда гормонов растений клетки начинают удлиняться только в одном направлении. Наиболее сильно выраженным действием такого рода обладают гиббереллины (гл. 12, разд. 3.1), способные вызывать очень быстрое удлинение клеток.
Удлинение клеток растений, несомненно, связано с разрывом старых и образованием новых связей между полисахаридами. Однако при этом сохраняется целостность целлюлозных волокон и их способность скользить друг относительно друга [114].
Клеточные стенки дрожжей и грибов состоят из глюканов, хитина и маннан-белкового комплекса. Некоторые сильно разветвленные маннановые цепи играют роль видоспецифичных антигенов [118]. Подобно антигенам поверхностей животных и бактериальных клеток, антигены растительных клеток характеризуются огромным структурным многообразием, что имеет важное значение для медицины. Удобным объектом для изучения генетических аспектов биосинтеза ферментов, участвующих в синтезе маннанов, являются дрожжи. Их можно выращивать как в гаплоидных, так и в гибридно-диплоидных формах, что значительно облегчает генетический анализ.
Вопросы и задачи
1. Стеариновую кислоту (1,16 г) растворили в 100 мл этанола. 10 мкл полученного раствора нанесли пипеткой на чистую поверхность разбавленного раствора НСl (в неглубоком лотке), так, чтобы образовался монослой стеариновой кислоты. Этот слой сжимали (путем перемещения тефлоновой пластинки) до тех пор, пока поверхностное давление п, резко возрастая, не достигало значения 20 дин∙см-1. Заметим, что п = у0—у, где у — поверхностное натяжение, измеряемое при наличии пленки, а у0 — более высокое поверхностное натяжение чистой воды. Сжатая пленка покрыла площадь 20X24 см. Рассчитайте поперечное сечение алкильной цепи в стеариновой кислоте. (См. Davenport J. В, in: Biochemistry and Methodology of Lipids (Johnson A. S., Davenport J. B, eds.), pp. 47—83. Wiley-Interscience, New York, 1971; and Phillips M. C, in: Progress in Surface and Membrane Science (Damelli J. F.), Rosenberg D. M., Cadenhead D. A, eds.), Vol. 5, pp. 139—221, Academic Press, New York, 1972.
2. В 1925 г. Э. Гортер и Ф. Грендел (J. Exp. Med, 41, 439) описали эксперименты, в которых они экстрагировали ацетоном липиды из мембран эритроцитов и получили из них монослой, после чего измеряли площадь сдавленного монослоя. Далее они оценили площадь поверхности эритроцитов и рассчитали, что отношение площади липидов (в виде монослоя) к площади поверхности эритроцита составляет 1,9—2,0. В ходе более поздних экспериментов [11] было установлено, что в каждой мембране эритроцита содержится 4,5∙10-16 моль фосфолипидов и 3,1∙10-16 моль холестерина.
а. Какую площадь будет занимать монослой, если принять, что поперечные сечения молекул фосфолипида и холестерина в мембране равны соответственно 0,70 и 0,38 нм2?
б. Чему будет равно отношение площади, рассчитанной в а), к площади поверхности эритроцита, если измеренная площадь его поверхности равна 167 мкм2?
в. Как вы объясните разницу между полученной величиной и соответствующей величиной, приведенной в работе Гортера и Грендела? (см. [11]).
3. Ниже приведены экспериментальные данные, полученные при изучении строения и функции биологических мембран. Объясните связь приведенных данных со строением мембран.
а. Многие макроциклические антибиотики (нонактин, валиномицин и др.) с высокой степенью избирательности связываются с катионами щелочных металлов, образуя комплексы с отношением 1 : 1. Эти комплексы хорошо растворимы в неполярных органических растворителях. Указанные антибиотики повышают электропроводность и проницаемость искусственных фосфолипидных мембран для катионов щелочных металлов. Валиномицин повышает электропроводность тилакоидных мембран хлоропластов в присутствии ионов К+, но не Na+; он также разобщает окислительное фосфорилирование в митохондриях (см. гл. 10).
б. При обработке интактных хлоропластов галактолипазой из галактозилдиглицеридов освобождается галактоза. При обработке теней эритроцитов фосфолипазой С освобождается около 75% липидного фосфора в водорастворимой форме. Ни в одном из случаев структурная целостность мембраны не нарушается.
в. Используя в качестве гаптена сфингомиелин, можно получить специфические антитела к этому липиду. Можно показать, что антитела взаимодействуют с тенями эритроцитов, но только с одной из сторон мембраны.
4. Опишите строение биологических мембран и специфические функции липид-, белок- и углевод-содержащих компонентов. В чем состоят различия между внутренней и наружной поверхностями мембраны?
5. Сравните распределение триглицеридов, фосфатидилхолина, фосфатидилэтаноламина, сфингомиелина, гликолипидов и холестерина внутри клеток. Оцените различия между разными сторонами мембран.
6. На каких химических принципах основана модификация компонентов клеточной поверхности с использованием в качестве реагентов следующих соединений: а) лактопероксидазы, б) галактозооксидазы, в) формилметионилсульфонметилфосфата, г) диазониевой соли дииодосульфаниловой кислоты, д) флуоресцирующих антител, е) антител, связанных с ферритином. Напишите уравнения соответствующих химических реакций. Укажите, какие поверхностные группы модифицируются. Перечислите преимущества указанных реагентов.
7. Какой из детергентов будет более эффективен в диапазоне pH от 2 до 3 — лаурилсульфонат натрия или лаурат натрия? Почему?
8. Какое количество энергии потребуется для транспорта 3 эквивалентов Na+ из клетки в среду и 2 эквивалентов К+ из внешней среды внутрь клетки, если в клетке концентрации этих ионов равны соответственно 10 мМ и 100 мМ, а в жидкой среде, в которой находятся клетки, 100 мМ и 5 мМ? Сравните полученную величину с ∆G' гидролиза АТР при pH 7. Считайте, что мембрана проницаема для ионов Сl-.
9. Считают, что клетка Е. coli содержит приблизительно 105 молекул белка оболочки, мол. вес которого 36 500. Какая часть поверхности бактерии будет покрыта белком, если молекулы белка имеют форму сфер, причем эти сферы плотно упакованы в гексагональные структуры? Каков диаметр молекулы белка? Какой должна быть площадь белковой молекулы, если 105 молекул покрывают полностью всю поверхность? Предложите форму белковой молекулы, удовлетворяющую этому требованию.
10. Проследите за перемещением ионов Са2+ внутри клетки, благодаря которым запускается процесс сокращения миофибрилл начиная с момента, когда нервный импульс достигает мышечной клетки (в скелетной мышце млекопитающего), и кончая завершением процесса сокращения. Назовите специфические белки и (или) структуры, участвующие в рассматриваемых процессах.