Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Коферменты - особые природные специализированные реагенты
Тетрагидрофолиевая кислота и другие птериновые коферменты
Одноуглеродные группы и соединения в процессах метаболизма
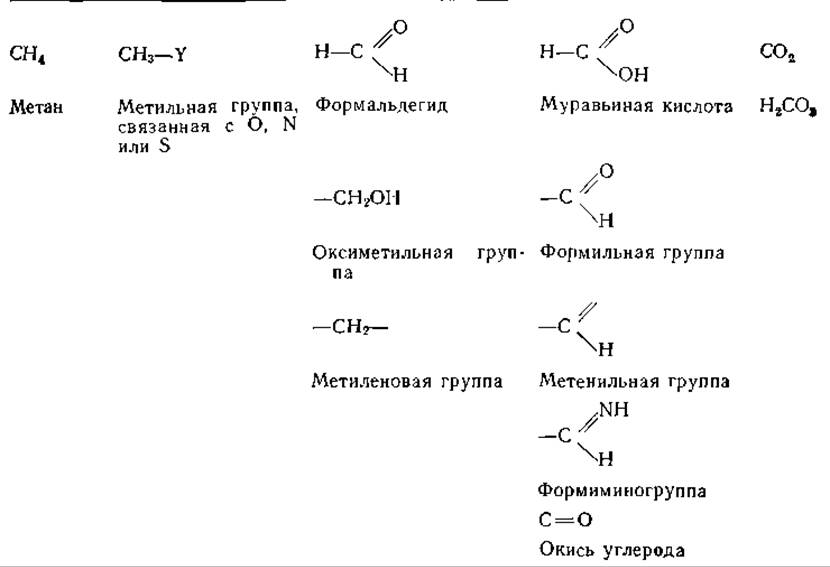
Некоторые одноуглеродные соединения и группы, играющие важную роль в метаболизме, приведены в табл. 8-5. Носителями одноуглеродных фрагментов, соответствующих трем различным уровням окисления (муравьиной кислоты, формальдегида и метильной группы), являются коферментные формы тетрагидрофолиевой кислоты. Хотя полностью восстановленное С1-соединение метан не может находиться в связанной форме, его биосинтез является Н4Fоl-зависимым процессом, так же как и биосинтез окиси углерода. На рис. 8-20 суммированы известные метаболические взаимоотношения соединений и групп, приведенных в табл. 8-5.
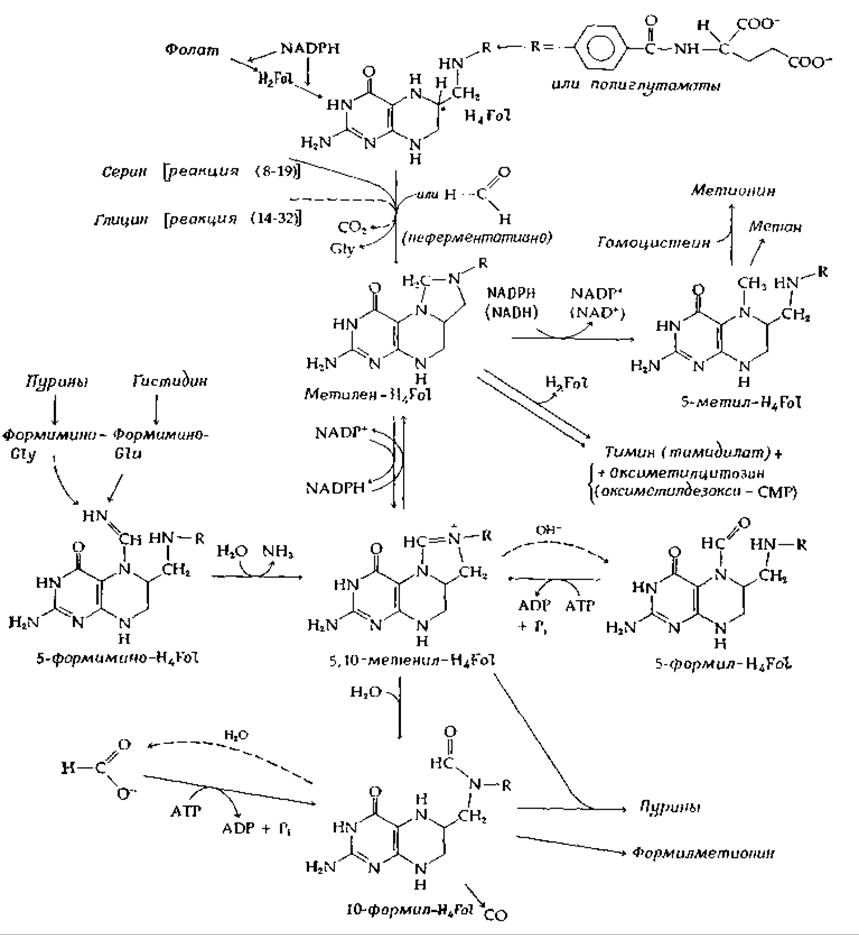
РИС. 8-20. Тетрагидрофолиевая кислота и ее одноуглеродные производные.
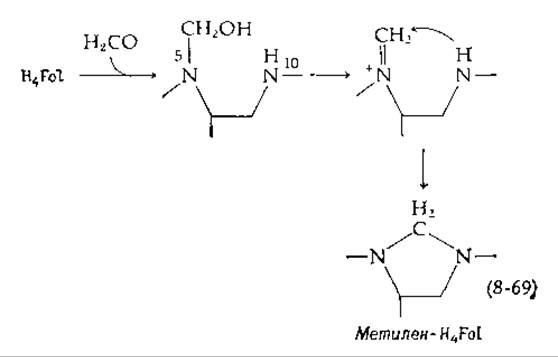
Во многих организмах основным предшественником С1-остатков служит серии. ß-Углерод серина отделяется в форме формальдегида путем прямого переноса на тетрагидрофолат с образованием метилен-H4Fol и глицина [уравнение (8-19)]. Последний в свою очередь может дать другую одноуглеродную единицу с потерей СО2 под действием H4Fol- и PLP-зависимой глициндекарбоксилазной системы [реакция (14-32)]. Свободный формальдегид также может присоединяться к H4Fol с образованием метилен-H4Fol [уравнение (8-69)] [155].
Таблица 8-5 Одноуглеродные соединения в порядке уровней окисления
В некоторых организмах, особенно в бактериях, которые способны использовать формиат в качестве единственного источника углерода, формиат вначале присоединяется к H4Fol с образованием 10-формил-Н4Fоl (нижний левый угол рис. 8-21).
Присоединенный к H4Fol одноуглеродный остаток формальдегидного уровня может быть либо окислен до 5,10-метенил-Н4Fоl и 10-формил-H4Fol, либо восстановлен до 5-метил-H4Fol. Организмы, утилизирующие формиат, могут соответственно восстанавливать 10-формил-Н4Fоl до метилен-Н4Fоl и переносить одноуглеродный остаток на глицин с образованием серина.
Роль 5-формил-Н4Fоl (лейковории или цитроворум-фактор)1) в метаболизме изучена хуже. Она может проявляться в переносе формильной группы от формилглутамата; существует фермент, превращающий 5-формил-Н4Fоl в 10-метенил-Н4Fоl с параллельным расщеплением АТР. Это соединение используется своеобразным способом при лечении (Некоторых особо злокачественных опухолей. Вслед за хирургическим удалением опухоли больному периодически вводят метотрексат в дозах, обычно смертельных. Затем, спустя 36 ч, больного спасают инъекцией 5-формил-Н4Fоl. Механизм «спасающего» действия 5-формил-Н4Fоl раскрыт не до конца.
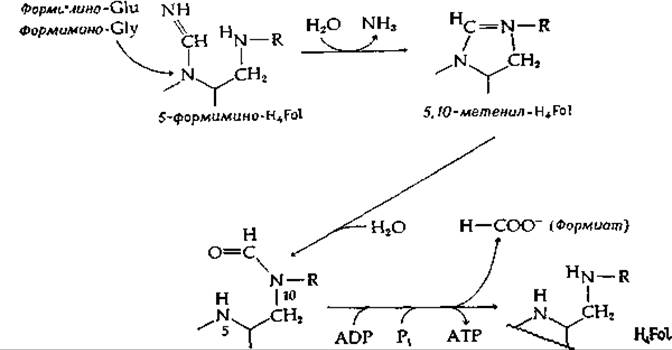
РИС. 8-21. Реакция, обеспечивающая фосфорилирование на субстратном уровне при посредстве 10-формил-Н4Fоl у бактерий вида Clostridium.
Другим метаболическим источником одноуглеродных фрагментов является диссимиляция гистидина, которая происходит как у бактерий, так и у животных, через формиминоглутаминовую кислоту. Последняя переносит группу —CH=NH на H4Fol с образованием 5-формимино-H4Fol, которая в свою очередь превращается в 5-10-метенил-Н4Fоl и аммиак. У бактерий, сбраживающих пурины, промежуточным соединением является формиминоглицин. И здесь формиминогруппа переносится на H4Fol и дезаминируется, переходя в 10-формил-Н4Fоl. Фермент, катализирующий превращение формиата в 10-формил-Н4Fоl, имеет очень высокую активность у этих организмов [158]; полагают, что он функционирует в обратном направлении, обеспечивая синтез АТР при этом типе брожения (рис. 8-21). В качестве промежуточного соединения образуется, возможно, формилфосфат [158а].
Как 5-10-метенил-Н4Fоl, так и 10-формил-Н4Fоl выступают в роли агентов биологического формилирования, и обе они необходимы при синтезе пуринов (гл. 14, разд. Л,3). Примечательная реакция происходит в газовой железе физалии, которая вырабатывает большие количества окиси углерода, по-видимому, из 10-формил-Н4Fоl. Окись углерода возникает также в природных условиях при метаболизме бактерий. Этот аспект экологической биохимии весьма актуален в связи с поступлением в биосферу больших количеств СО в результате деятельности человека: бактерии производят его несравненно больше, чем человек.
Метилен-H4Fol выступает в роли непосредственного предшественника 5-метильной группы тимина, а также оксиметильной группы оксиметилцитозина. В ходе образования тимина кофермент окисляется в дигидрофолиевую кислоту [реакция (14-51)], которая для завершения каталитического цикла должна быть восстановлена дигидрофолатредуктазой. Восстановление метилен-Н4Fоl до 5-метил-H4Fоl служит одним из источников метана в метаболизме бактерий (разд. М, 8) и у всех живых организмов обеспечивает поставку метальных групп, необходимых в больших количествах для синтеза метионина и модификации белков, нуклеиновых кислот и других биохимических соединений, осуществляемых путем метилирования специфических групп.
1) В 1949 г. было установлено, что N5-формил-Н4Fоl является фактором роста Leuconostoc citrovorum, откуда и происходит название этого соединения цитроворум-фактор.