Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Организация метаболизма: катаболические пути
Цикл трикарбоновых кислот
Регуляция цикла
Какими же факторами определяется скорость функционирования цикла трикарбоновых кислот? Как и в других важнейших метаболических путях, работает несколько разных механизмов контроля, причем в различных условиях скорость лимитируется разными стадиями процесса [18] Главными факторами являются: 1) скорость поступления ацетильных групп (которая в свою очередь может зависеть от наличия свободного неацилированного СоА); 2) наличие оксалоацетата и 3) скорость реокисления NADH в NAD+ в цепи переноса электронов (гл. 10). Обратите внимание (рис. 9-3), что ацетил-СоА служит положительным эффектором для превращения пирувата в оксалоацетат. Таким образом, ацетил-СоА «включает» процесс образования соединения, требующегося для его собственного метаболизма. В отсутствие пирувата функционирование цикла может затормозиться из-за недостатка оксалоацетата. По-видимому, именно так и происходит в тех случаях, когда в печени метаболизируются высокие концентрации этанола Последний окисляется в ацетат, но не может превратиться в оксалоацетат. Накапливающиеся ацетильные группы превращаются в кетоновые тела, которые, однако, медленно окисляются в цикле. Аналогичная проблема возникает при метаболизме жирных кислот в условиях нарушения углеводного обмена, например в случае диабета (дополнение 11-В).
Скорость окислительных стадий цикла определяется скоростью реокисления NADH в цепи переноса электронов. При некоторых условиях ее может лимитировать скорость поступления О2. Однако в аэробных организмах она обычно определяется концентрацией ADP и (или) Рі, доступных для превращения в АТР в процессе «окислительного фосфорилирования» (гл. 10). Если в ходе катаболизма образуется больше АТР, чем это необходимо для энергетических потребностей клетки, концентрация ADP падает до низкого уровня, выключая, таким образом, процесс фосфорилирования. Одновременно АТР, присутствующий в высоких концентрациях, действуя по принципу обратной связи, ингибирует процессы катаболизма углеводов и жиров. Это ингибирование осуществляется во многих пунктах метаболизма, часть которых показана на рис 9-3. Важным участком, на котором осуществляется такое ингибирование, является пируватдегидрогеназный комплекс (гл 8, разд К2) [19]. Другим таким участком служит цитратсинтетаза — фермент, катализирующий первую реакцию цикла трикарбоновых кислот [20]. Правда, существуют сомнения относительно того, имеет ли такое ингибирование физиологическое значение [16]. Уровень фосфорилирования аденилатной системы может регулировать работу цикла еще и другим способом, связанным с потребностью в GDP на стадии е цикла (рис. 9-2). В митохондриях GTP в основном используется для превращения АМР в ADP. Следовательно, образование GDP зависит от АМР — соединения, которое образуется в митохондриях при использовании АТР для активации жирных кислот [уравнение (9-1)].
В некоторых бактериях (включая Е. coli) АТР не оказывает ингибирующего действия на цитратсинтетазу, но такое действие оказывает NADH; в этом случае контроль определяется не уровнем фосфорилирования адениннуклеотидной системы, а окислительно-восстановительным потенциалом NAD+-системы [21]. Активность сукцинатдегидрогеназы может зависеть от окислительно-восстановительного состояния убихинона (гл. 10, разд. Г).
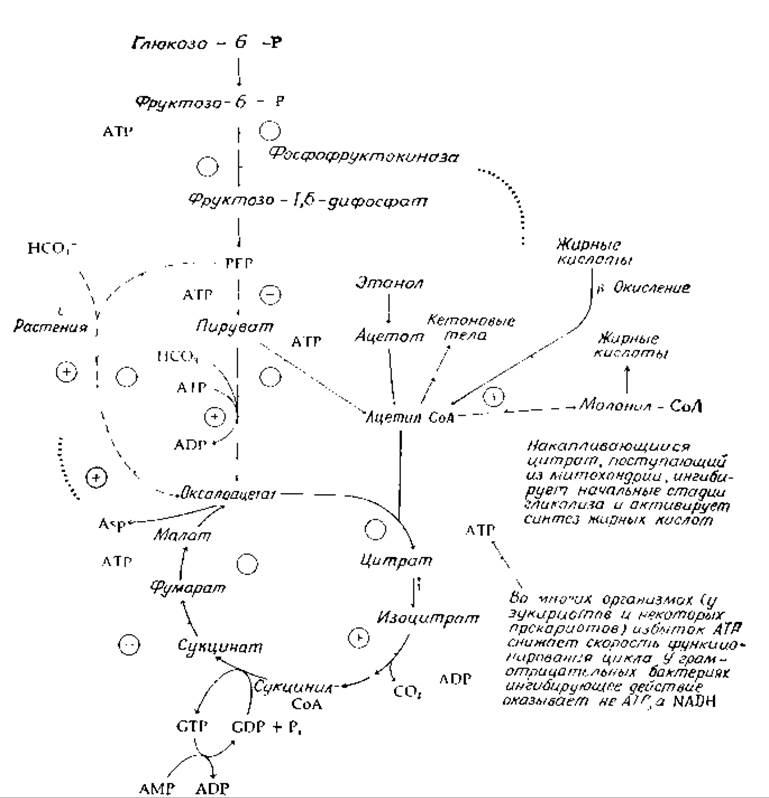
РИС 9-3 Схема регуляции гликолиза и цикла трикарбоновых кислот (см также рис 11-11)
Вторым сигналом, оповещающим клетку о слишком высокой скорости процессов катаболизма, является выход из митохондрий накопившегося цитрата. Это имеет существенное значение для синтеза жирных кислот; одновременно цитрат действует как отрицательный эффектор на фосфофруктокиназу (рис. 9-3) и таким образом снижает общую скорость гликолиза. При понижении концентрации АТР и накоплении ADP последний служит положительным эффектором для окисления изоцитрата, вызывая в результате быстрое снижение концентрации цитрата.
Нетрудно понять, что возможны и многие другие способы регуляции. Можно представить, например, регуляцию, зависящую от концентраций Pi и СоА. Существует точка зрения, согласно которой может регулироваться активность каждого из ферментов цикла и при соответствующих условиях начинает действовать тот или иной механизм регуляции [21].