Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Организация метаболизма: катаболические пути
Цикл трикарбоновых кислот
Катаболизм промежуточных продуктов цикла трикарбоновых кислот
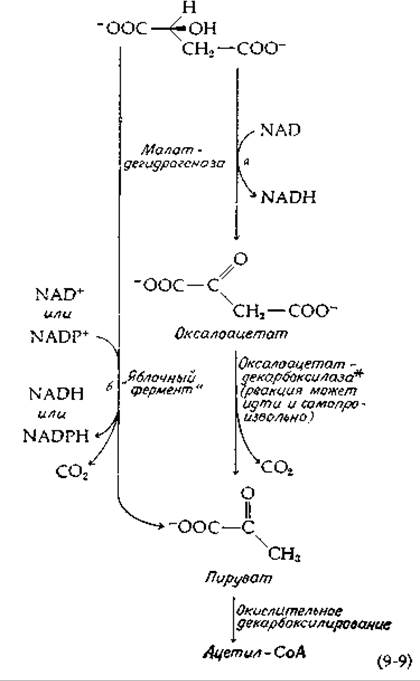
Иногда возникает необходимость в том, чтобы в клетке окислялись большие количества какого-то одного из компонентов цикла трикарбоновых кислот [22]. Так, бактерии, использующие в качестве источника углерода сукцинат, должны окислять его для обеспечения энергией, а также затрачивать какую-то часть сукцината на образование углеводов, липидов и других материалов. Полное окисление любого промежуточного продукта цикла трикарбоновых кислот может быть осуществлено путем превращения его в малат, после чего следует либо окисление малата в оксалоацетат [уравнение (9-9, стадия а)] и далее декарбоксилирование оксалоацетата в пируват (ß-расщепление), либо окисление и декарбоксилирование малата до пирувата [уравнение (9-9, стадия б)] с помощью одного «яблочного» фермента [уравнение (7-75)] без промежуточного образования свободного оксалоацетата. Путь б, по-видимому, наиболее важен. В митохондриях животных имеется два «яблочных» фермента, один из которых специфичен к NADP+, а другой может функционировать также и с NAD+ [23]. Оба фермента обладают сложными регуляторными свойствами. Например, NADP+-зaвисимый фермент активируется высокими концентрациями свободного СоА и ингибируется NADH. На этот счет предлагается следующее объяснение: при замедлении процессов гликолиза сильно повышается концентрация свободного СоА, который «включает» процесс окисления малата [24]. С другой стороны, быстрый гликолиз приводит к повышению концентрации NADH и к ингибированию «яблочного» фермента. В результате возрастает концентрация оксалоацетата и повышается активность цикла трикарбоновых кислот. Полагают также, что «яблочный» фермент функционирует в цикле, приводящем к образованию NADPH [уравнение (11-13)].