Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Организация метаболизма: катаболические пути
Катаболизм сахаров
Гликолиз
Поскольку у большинства сахаров при каждом из атомов углерода имеется атом кислорода, химическая атака, приводящая к окислению, оказывается возможной в любой точке молекулы. Каждый сахар содержит потенциально свободную альдегидную или кетонную группу, и карбонильная группа может быть легко перемещена в соседнее положение с помощью изомераз. Альдольное расщепление также оказывается возможным во многих точках. По этим причинам метаболизм углеводов сложен и разнообразен. Однако в энергетике большинства организмов, в том числе и человека, гликолитический1) путь, с помощью которого гексозы превращаются в пируват (гл. 7, разд. А,5, и рис. 9-7), намного опережает по значению все другие.
1) Слово «гликолиз» означает расщепление сахаров или гликогена. Первоначально оно относилось только к анаэробному брожению, завершающемуся образованием молочной кислоты или этанола и СО2, однако теперь имеет более широкое значение и используется для описания распада сахаров, проходящего через образование глюкозо-6-фосфата, фруктозодифосфата и пирувата как в присутствии кислорода, так и при его отсутствии.
Открытие гликолиза последовало непосредственно за экспериментами Бюхнера, а также Гардена и Ионга по сбраживанию сахара дрожжевым соком (гл. 8, разд. 3). Вскоре с изучением спиртового брожения слились исследования другого направления, связанные с изучением мышц. Физиологи заинтересовались процессом, благодаря которому изолированная мышца могла получать энергию для сокращения в отсутствие кислорода. Хилл показал, что энергию обеспечивает превращение гликогена в лактат, а несколько позднее Мейергоф продемонстрировал, что происходящие при этом химические реакции сходны с теми, которые наблюдаются при спиртовом брожении. Установление структуры и функции пиридиннуклеотидов в 1934 г. (гл. 8, разд. 3) совпало по времени с важными исследованиями по изучению гликолиза, проведенными Эмбденом во Франкфурте и Парнасом в Польше. Таким путем вскоре была выяснена последовательность реакций гликолиза (путь Эмбдена — Мейергофа—Парнаса). Все ферменты, катализирующие отдельные стадии процесса, к настоящему времени выделены, закристаллизованы и подробно изучены.
а. Образование пирувата
Превращение глюкозы в пируват требует участия десяти ферментов (рис. 9-7). Вся последовательность реакций может быть разбита на четыре стадии: подготовка к разрыву цепи (реакции 1—3), разрыв цепи и установление равновесия между триозофосфатами (реакции 4 и 5), окислительное образование АТР (реакции 6 и 7) и превращение 3-фосфоглицерата в пируват (реакции 8—10).
При подготовке к разрыву цепи свободная глюкоза фосфорилируется молекулой АТР при участии гексокиназы (реакция 1). В тканях имеется несколько гексокиназ; некоторые из них относительно неспецифичны и катализируют также фосфорилирование других сахаров, например маннозы и галактозы (гл. 6, разд. Е,2; гл. 7, разд. Д,6). Продукт фосфорилирования, глюкозо-6-фосфат, может образовываться и без затраты АТР путем отщепления глюкозильных остатков от гликогена, осуществляемого гликогенфосфорилазой (реакция 1а), с последующим действием фосфоглюкомутазы [реакция 16; см. также уравнение (7-26)]. Последний фермент переносит фосфорильную группу от кислорода при С-1 на кислород при С-6.
Почему клетки начинают метаболизм сахара с того, что присоединяют к нему фосфорильную группу? Во-первых, потому, что заряженная фосфорильная группа способствует связыванию сахарофосфата с ферментами. Во-вторых, с кинетической точки зрения выгодно запускать длинную последовательность реакций с практически необратимой реакцией, такой, как фосфорилирование глюкозы. Кроме того, предполагается возможность участия фосфатной группы в каталитическом процессе (гл. 7, разд. К, 2, а).
Реакция 2 на рис. 9-7 является простой изомеризацией, перемещающей карбонильную группу в положение С-2, в результате чего становится возможным ß-расщепление на два трехуглеродных фрагмента. Перед этим расщеплением происходит второе фосфорилирование (реакция 3), приводящее к образованию фруктозо-1,6-дифосфата. Тем самым после расщепления фруктозодифосфата альдолазой в каждой из двух образовавшихся половин оказывается фосфатная группа. Эта вторая «затравочная реакция» (реакция 3) является первой стадией последовательности, уникальной для гликолиза. Поэтому неудивительно, что катализирующий ее фермент фосфофруктокиназа — один из наиболее эффективно регулируемых катализаторов в гликолитической последовательности [уравнение (6-91); гл. 11, разд. Е,4].
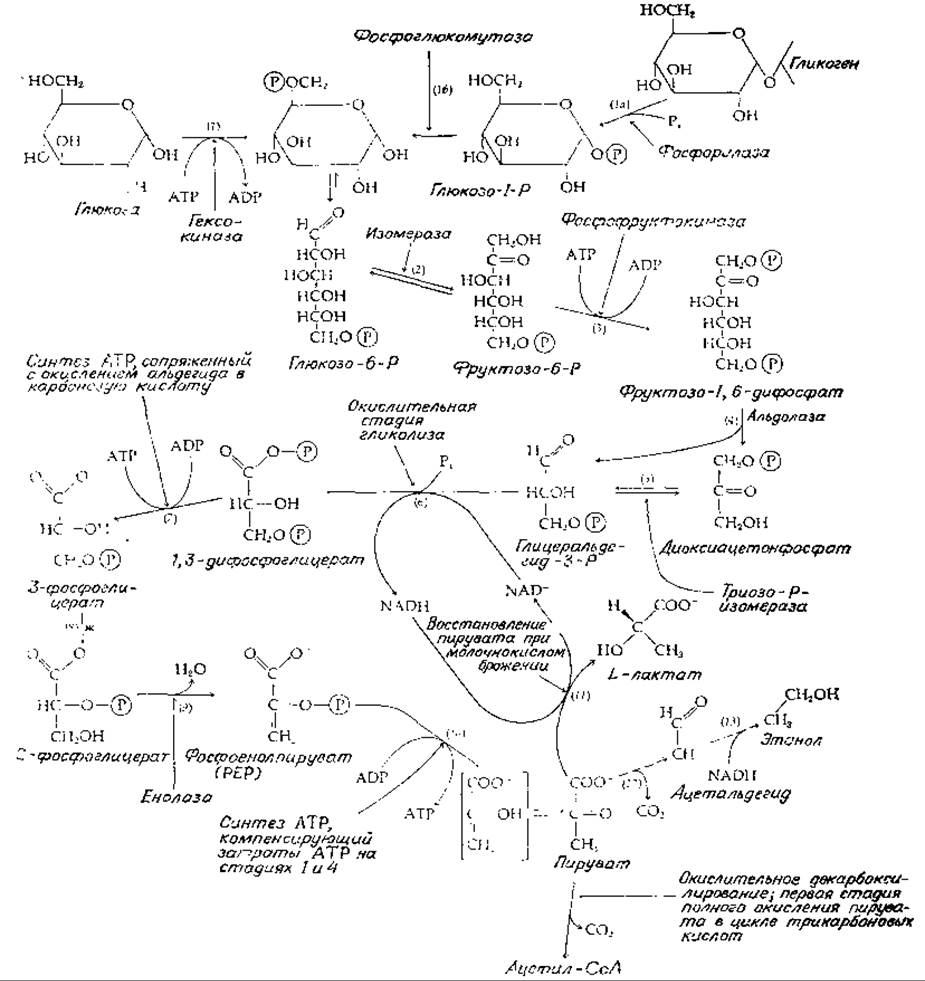
РИС. 9-7. Гликолиз, расщепление остатков гексозы (через стадию образования пирувата).
Расщепление фруктозодифосфата (реакция 4) катализируется альдолазой [уравнение (7-64)]; в результате образуются глицеральдегид-3-фосфат и диоксиацетонфосфат. Между этими двумя триозофосфатами в результате действия изомеразы устанавливается равновесие (реакция 5; см. также гл. 7, разд. И, 4). Таким образом, обмен обеих половинок гексозы может пойти по пути превращения в пируват через глицеральдегид-3-фосфат. В то же время для диоксиацетонфосфата существует и другой путь, связанный с восстановлением в глицерофосфат — предшественник липидов и промежуточный продукт в некоторых типах брожения.
Окисление глицеральдегид-3-фосфата в соответствующую карбоновую кислоту, 3-фосфоглицериновую кислоту (рис. 9-7, реакции 6 и 7), сопряжено с синтезом молекулы АТР из ADP и Рі(рис. 8-13). Отметим, что в процессе гликолиза на одну расщепленную молекулу гексозы образуются две молекулы АТР и одновременно две молекулы NAD+ превращаются в NADH.
Превращение 3-фосфоглицерата в пируват начинается с переноса фосфорильной группы от кислорода при С-3 на кислород при С-2 (реакция S) с последующим дегидратированием путем обычного а,β-элиминирования, катализируемого енолазой (реакция 9). Образовавшийся продукт фосфоенолпируват (РЕР; гл. 7, разд. К, 3, г) является «высокоэнергетическим» соединением, фосфорильная группа которого может быть легко перенесена на ADP (под действием фермента пируваткиназы); остающийся при этом енол пировиноградной кислоты (на рис. 9-7 он помещен в квадратные скобки) самопроизвольно превращается в значительно более устойчивый пируват-ион (сравните с уравнением 7-59). Поскольку на каждую молекулу глюкозы образуются две молекулы РЕР, этот процесс восполняет затрату двух молекул АТР, происходящую на начальных стадиях образования фруктозо-1,6-дифосфата из глюкозы.
б. Дальнейший метаболизм пирувата
При аэробном метаболизме, происходящем в большинстве тканей нашего организма, пируват путем окислительного декарбоксилирования превращается в ацетил-СоА, который далее подвергается полному окислению в цикле трикарбоновых кислот (рис. 9-2). NADH, образовавшийся в реакции 6, а также при окислительном декарбоксилировании пирувата и в последующих реакциях цикла трикарбоновых кислот, далее вновь окисляется в цепи переноса электронов в митохондриях. Другой важный путь превращения пирувата связан с брожением. Например, фермент лактатдегидрогеназа катализирует восстановление пирувата за счет NADH (реакция 11) в L-лактат (у многих бактерий образуется D-лактат). Как показано на рис. 9-7, эта реакция может быть сопряжена с реакцией 6, в которой образуется NADH. В результате может происходить сбалансированный процесс сбраживания глюкозы в лактат в отсутствие кислорода (гл. 7, разд. А, 6). В сходном процессе дрожжевые клетки вначале декарбоксилируют пируват (а-расщепление) в ацетальдегид, который затем восстанавливается в этанол, используя NADH, образовавшийся в реакции 6 (рис. 9-7, реакции 12 и 13). Эти реакции брожения будут далее рассматриваться в разд. Е.