Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Организация метаболизма: катаболические пути
Брожение: «жизнь без кислорода»
Смешанное кислое брожение
Кишечные бактерии, в том числе Е. coli, превращают глюкозу в этанол, уксусную кислоту и муравьиную кислоту (последняя может далее распадаться на СO2 и Н2). Стехиометрия может быть разной, однако в идеализированном виде этот процесс брожения можно описать уравнением
![]()
Детали процесса и окислительно-восстановительный баланс иллюстрирует уравнение
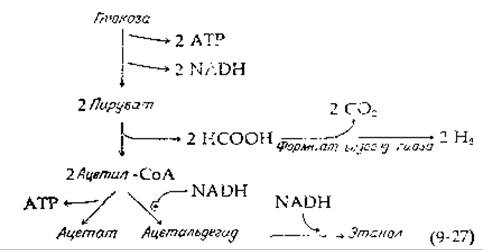
Сначала глюкоза превращается в пируват с образованием двух молекул АТР и двух молекул NADH. Пируват расщепляется далее в результате пируват-формиат-лиазной реакции (гл. 8, разд. К, 5). Продуктами являются ацетил-СоА и муравьиная кислота [уравнение (9-27); рис. 9-9, путь д].
Половина молекул ацетил-СоА расщепляется до ацетата через ацетилфосфат с образованием АТР. Другая половина молекул восстанавливается в две стадии в этанол, используя две молекулы NADH, образовавшиеся ранее при окислении триозофосфата [уравнение (9-27)]. Общий энергетический выход процесса составляет три молекулы АТР на одну молекулу глюкозы. «Эффективность» получается равной 3∙34,5/225=46%. Часть глюкозы превращается также и в D-молочную и янтарную кислоты (рис. 9-9, е), отсюда и название смешанное кислое брожение.
В некоторых вариантах смешанного кислого брожения, как это наблюдается, например, у бактерий Shigella, муравьиная кислота накапливается, однако в других случаях (в частности, у Е. coli) муравьиная кислота превращается в СО2 и Н2 [уравнение (9-27)]. Равновесие в этой реакции поддерживает формиат-водород-лиазная система, состоящая из двух ферментов. Селенсодержащая формиатдегидрогеназа (разд. В, 3) катализирует окисление формиата в СО2 с участием NAD+, тогда как связанная с мембраной гидрогеназа, вероятно представляющая собой железо-серный белок (гл. 10, разд. Е, 1,а), устанавливает равновесие в реакции NADH+H+→NAD++H2. Гидрогеназа служит также для освобождения Н2 при избыточных количествах NАDH. Как указал Кребс, избыточное накопление NADH в процессе роста клеток может быть обусловлено необходимостью осуществлять биосинтез сразу многих компонентов, в частности аминокислот. В тех случаях, когда глюкоза является единственным источником углерода, наблюдается преобладание окислительных стадий над восстановительными [38]. Избыток восстановительных эквивалентов может быть удален в виде Н2 или может быть использован для образования в высокой степени восстановленных продуктов, таких, как сукцинат.
В бактериях рода Aerobacter и Serratia часть образующегося пирувата конденсируется (с одновременным декарбоксилированием) в ацетолактат (являющийся также промежуточным продуктом в биосинтезе валина; рис. 14-10), который в свою очередь декарбоксилируется в ацетоин [уравнение (9-28); рис. 9-9, путь д]. Ацетоин восстанавливается за счет NADH в 2,3-бутандиол. При этом третья молекула пирувата превращается в этанол, водород и СО2 [уравнение (9-28)]. В итоге получается следующее суммарное уравнение
![]()
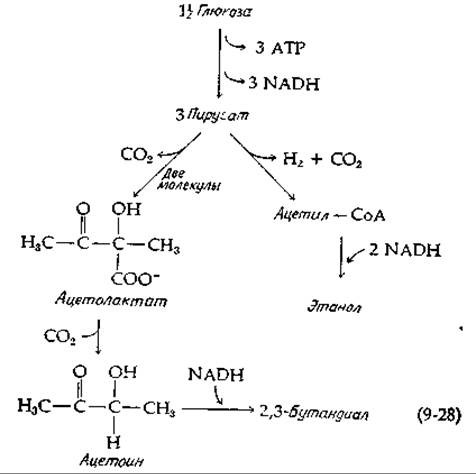
Эти же организмы образуют и кислоту в результате реакций, которые рассматривались выше. Реакция (9-29) лежит в основе промышленного получения бутандиола (который затем может быть неферментативным путем дегидрирован в бутадиен).
Смешанное кислое брожение встречается не только у бактерий. Так, трихомонады, паразитические жгутиковые, относящиеся к типу простейших, тоже способны в анаэробных условиях превращать пируват в ацетат, сукцинат, СО2 и Н2. У этих организмов нет митохондрий, но имеются напоминающие микротельца частицы, названные гидрогеносомами, способные превращать пируват в ацетат, СО2 и Н2 [39]. Фермент, катализирующий расщепление пирувата, по-видимому, не содержит липоата и, возможно, близок по свойствам пируват: ферредоксин—оксидоредуктазе клостридий [уравнение (8-66)]. В гидрогеносомах находится также активная гидрогеназа.
Многие беспозвоночные являются истинно факультативными анаэробами, способными выживать длительное, а иногда и неопределенно долгое время в отсутствие кислорода [39а, b]. К ним принадлежат черви Ascaris (рис. 1-10), а также устрицы и другие моллюски. К числу главных конечных продуктов их анаэробного метаболизма относятся сукцинат и аланин. Первый может образовываться при смешанном кислом брожении наряду с пируватом. Далее у Ascaris lumbricoides, который фактически является облигатным анаэробом, пируват превращается в ацетат. Однако у моллюсков пируват вступает в реакцию переаминирования с глутаматом с образованием аланина и а-кетоглутарата; из кетоглутарата в результате окислительного декарбоксилирования может образоваться сукцинат. Для этих реакций, очевидно, необходимо наличие глутамата или некоторых других аминокислот (например, аргинина), которые могут превращаться в глутамат.