Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Биосинтез; как образуются новые молекулы
Специальные аспекты метаболизма жирных кислот
Ненасыщенные жирные кислоты и продукты их превращения
Жирные кислоты с одной или более ненасыщенными двойными связями придают клеточным мембранам необходимую текучесть, а также служат предшественниками других компонентов клетки. Имеются значительные различия в способе введения двойной связи в жирные кислоты у животных, простейших, грибов и некоторых бактерий, с одной стороны, и бактерий типа Е. coli, способных жить в анаэробных условиях, — с другой. У последней основным представителем ненасыщенных жирных кислот является вакценовая кислота; она образуется путем удлинения цепи после введения цис-двойной связи на стадии синтеза С10-фрагмента. Под действием ß-оксидеканоилтиоэфирдегидратазы, присутствующей в клетках Е. coli. происходит отщепление ß-гидроксила с образованием главным образом цис-β,у- (а не транс-а,ß-) ненасыщенного соединения [уравнение (12-14)]. По механизму этот процесс сходен с действием еноилгидратазы [уравнение (7-42)] в обратном направлении, причем заключенный в скобки транс-а,ß-ненасыщенный продукт (связанный с ферментом) изомеризуется в цис-β,у-ненасыщенное соединение посредством аллильной перегруппировки. Далее происходит удлинение указанного соединения до С16- и C18-производных СоА, т. е. соответственно в пальмитоил- и вакционил-СоА (рис. 12-6).
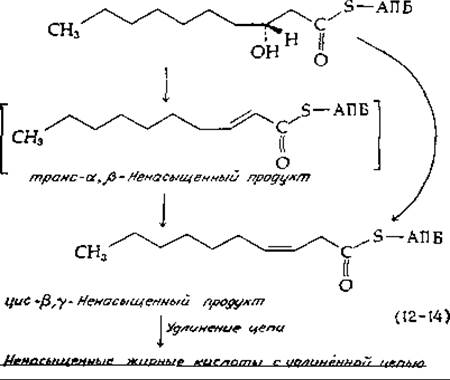
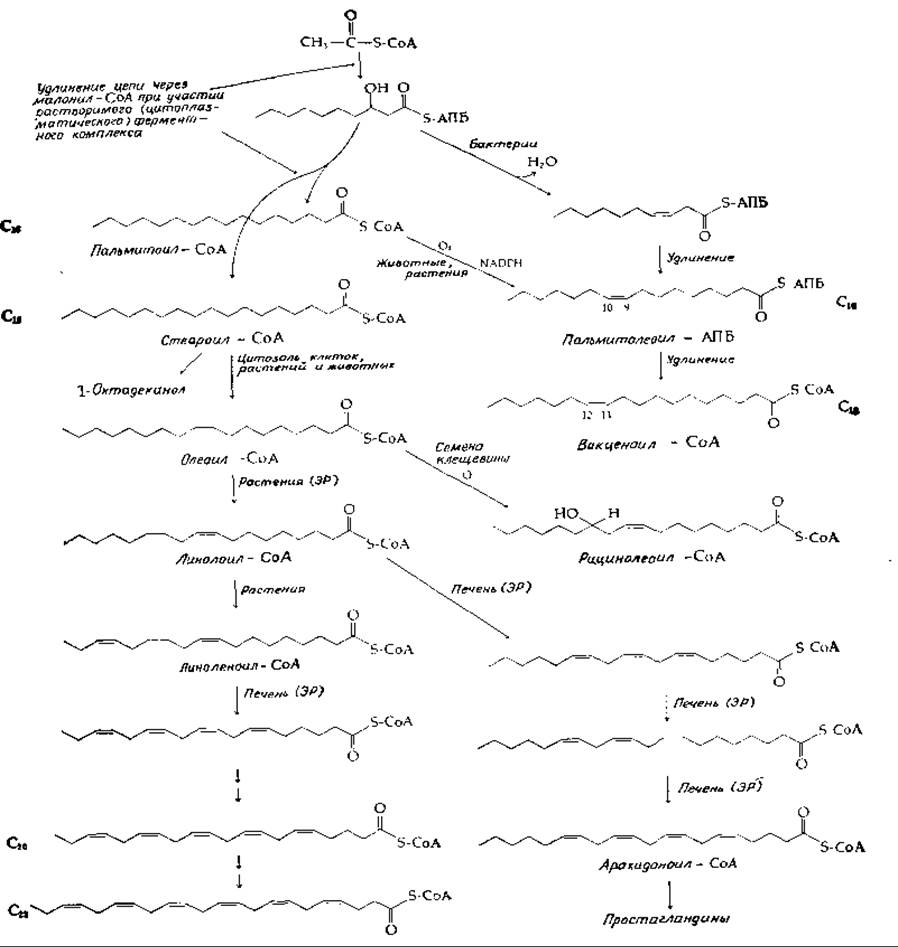
РИС. 12-6. Некоторые реакции биосинтеза жирных кислот.
Однако реакция дегидратации ß-оксидеканоил-АПБ1) составляет одно из ответвлений от основных путей биосинтеза [41], тогда как транс-а,ß-ненасыщенные длинноцепочечные ацильные соединения лежат на основном пути удлинения цепи до пальмитил-СоА (рис. 12-6, левая сторона).
1) Хотя на рис. 12-6 показаны производные как СоА, так и АПБ, однако многие этапы процесса пропущены. Нередко процесс биосинтеза начинается, когда жирная кислота присоединена к ацилпереносящему белку, но конечный продукт освобождается в форме СоА-производного (гл. 11, разд Г,6).
У высших растений, животных, простейших и грибов насыщенные жирные кислоты подвергаются действию десатураз (гл. 10, разд. Ж, 3), приводящих к образованию двойных связей, как правило, цис-конфигурации. Как у животных, так и у растений введение в насыщенную жирную кислоту первой двойной связи протекает в цитозоле. Образующийся при этом олеилкофермент в ходе реакций, изображенных на рис. 12-6, превращается в СоА-производные линолевой, линоленовой, арахидоновой и других полиеновых кислот. В клетках растений процессы десатурации протекают в эндоплазматическом ретикулуме при участии NADPH, генерируемого на свету ферредоксина и О2.
У животных превращения олеил-СоА в линолил-СоА не происходит. Вследствие этого полиненасыщенные жирные кислоты, такие, как линолевая, линоленовая и С20-арахидоновая, являются незаменимыми компонентами пищи. При отсутствии этих незаменимых жирных кислот растительного происхождения1) у животных затормаживается рост, возникают поражения кожи, повреждения почек, нарушается функция размножения. В настоящее время установлено, что одной, хотя, вероятно, не единственной, существенной функцией незаменимых жирных кислот является участие в синтезе (в качестве предшественников) «гормонов местного действия», а именно простагландинов (разд. Д, 3) [42]. Установлена особая роль арахидоновой кислоты в тромбоцитах, где под действием липоксигеназы из нее образуется 12-L-окси-5,8,10,14-эйкозатетраеновая кислота — фактор хемотаксиса нейтрофилов (дополнение 5-Ж).
На рис. 12-6 показан также процесс образования рицинолевой кислоты, специфического компонента семян клещевины и касторового масла (рис. 2-32). Некоторые организмы содержат циклопропановые жирные кислоты (рис. 2-32) [43]. Донором дополнительного углерода циклопропанового кольца служит S-аденозилметионин (SAM), причем углерод присоединяется по двойной связи в ацильной группе жирной кислоты, находящейся в составе фосфатидилэтаноламина — постоянного компонента мембран [уравнение (12-15)] [44,45].
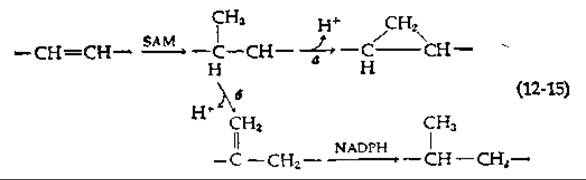
Из одного и того же карбоний-иона может образовываться либо циклопропановая жирная кислота [уравнение (12-15), реакция а], либо метенильное производное жирной кислоты [уравнение (12-15), реакция б]; последнее в свою очередь может в результате превратиться в жирную кислоту с разветвленной цепью. Указанный процесс представляет собой путь образования метилированных жирных кислот у некоторых бактерий [44].
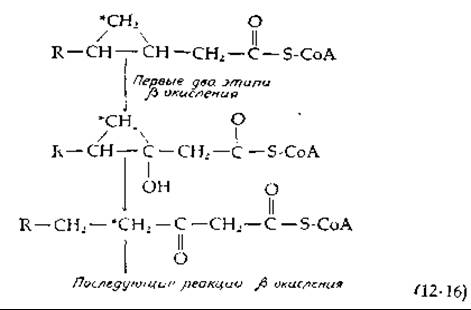
Циклопропановые жирные кислоты распадаются путем ß-окисления, который, однако, несколько видоизменяется, когда расщепление цепи достигает циклопропанового кольца [см. уравнение (12-16)] [46]. Раскрытие кольца у производных циклопропанола происходит очень легко, даже при умеренном неферментативном кислотно-щелочном катализе.
1) Арахидоновая кислота обычно не встречается у растений, а у животных образуется из линолевой кислоты, как показано на рис. 12-6.
В ненасыщенных жирных кислотах могут образоваться также ацетиленовые группы (—C ≡ C—). Это происходит, по-видимому, путем дегидрогенизации —СН = СН—, но ферменты, катализирующие процесс, мало исследованы. В качестве примеров природных ацетиленов назовем крепениновую кислоту (рис. 2-32), аллоксантин (разд. 3,3) и следующий любопытный углеводород, присутствующий в обычном васильке Centaurea cyanus [45]:
![]()