Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Биосинтез; как образуются новые молекулы
Стероидные соединения
Биосинтез стеринов
Большая часть стероидов в животном организме образуется из холестерина, который в свою очередь является производным сквалена1). Процесс превращения сквалена, протекающий в большинстве животных тканей, начинается с того, что под действием ферментной системы микросом в присутствии О2 и NADPH образуется 2,3-окись сквалена [91—94]. По-видимому, реакция проходит через стадию образования карбоний-иона при атаке протоном атома кислорода в эпоксидном кольце [уравнение (12-31)] [77]. Ток электронов (одновременный или поэтапный) приводит к замыканию всех четырех колец, а карбоний-ион остается у места присоединения боковой цепи к кольцу D [уравнение (12-31), этап а]. Перестройка структуры, ведущая к образованию ланостерина [уравнение (12-31), этап б], является примечательной реакцией, которая сопряжена с перемещением одного гидрид-иона и двух метальных групп, как показано стрелками в уравнении (12-31). Кроме того, при этом имеет место уход водорода в форме протона из положения С-9 Ланостерин1) используется в организме животных в качестве предшественника других стеринов. В растениях же, где холестерин отсутствует или содержится в очень небольшом количестве, основным предшественником при биосинтезе стеринов служит циклоартенол. Как показано в уравнении (12-31), этап в, для образования циклоартенола необходимо смещение протона (в виде гидрид-иона) и вытеснение им метильной группы у С-8. Отщепление протона от прилегающей метальной группы позволяет замкнуться циклопропановому кольцу.
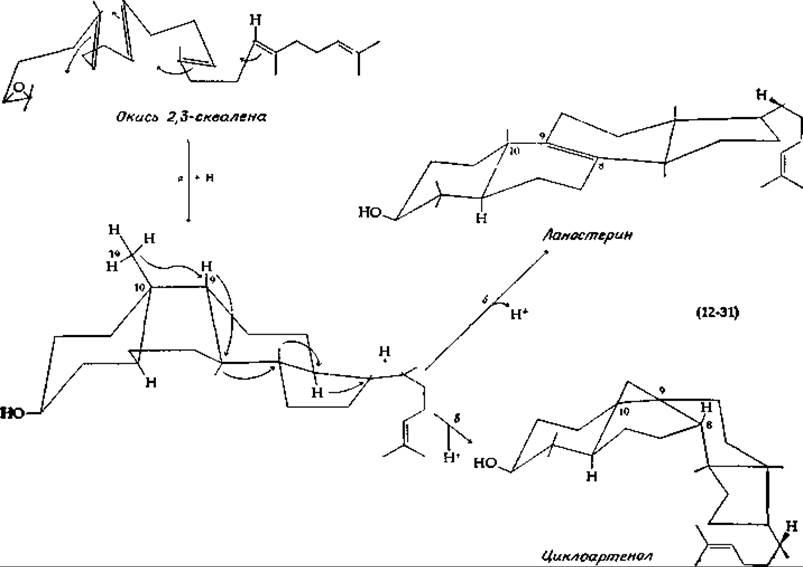
Превращение ланостерина в холестерин представляет собой сложный процесс, состоящий по крайней мере из 25 этапов. Многие из участвующих в процессе ферментов связаны с мембранами эндоплазматического ретикулума [95]. В процессе участвует также по крайней мере один растворенный в цитоплазме белок. Этот белок, транспортирующий стерин, функционирует как переносчик стерина от одного фермента к другому в ходе процесса превращения и, кроме того, влияет на реактивность присоединенной структуры [96, 97].
Как было установлено, удаление трех метальных групп ланостерина, миграция двойной связи в ß-кольце и насыщение двойной связи в боковой цепи могут осуществляться в ходе различных последовательностей реакций. Два варианта последовательностей показаны на рис. 12-15. Механизм реакции деметилирования в участке соединения колец С и D остается неясным, но аналогичные реакции отщепления метильных групп из положения С-4 кольца А изучены хорошо [95]. Каждая из метильных групп последовательно подвергается гидроксилированию при участии микросомной системы, сходной с цитохромом Р-450 (гл. 10, разд. Ж, 2, е) [97а], но использующей в качестве донора электронов не NADPH, a NADH. Окисление образующегося спирта в карбоновую кислоту и последующее превращение 3-ОН-группы в кетон способствует протеканию вслед за этим ß-декарбоксилирования.
1) Компонент ланолина, воскообразного жира овечьей шерсти, состоящего в основном из холестерина.
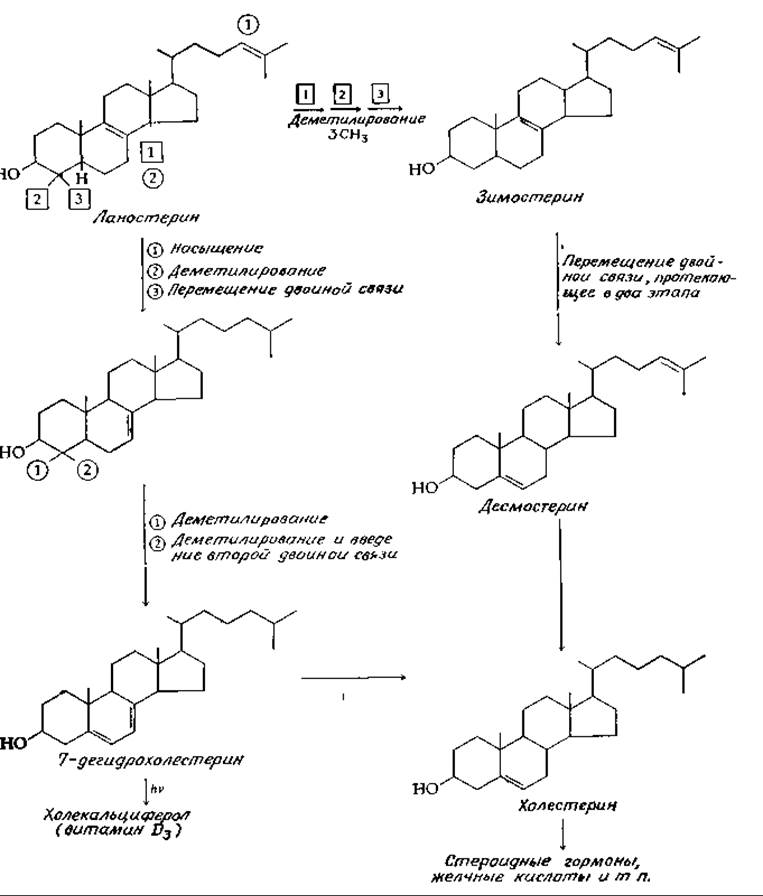
РИС 12-15 Превращение ланостерина в холестерин. Цифры в кружочках и квадратах показывают последовательность вовлечения в процесс определенных участков молекулы
К числу стеринов, образующихся у животных, относится также дегидрохолестерин — предшественник витамина D, содержащийся в значительном количестве в коже. В кишечном тракте под действием бактерий образуются ß-холестанол и его изомер ß-копростанол. Многие стерины растений отличаются от холестерина наличием дополнительных
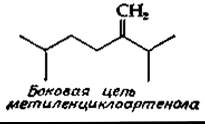
(одного или нескольких) атомов углерода в боковой цепи. Источником этих углеродных атомов служит S-аденозилметионин. Как уже упоминалось, стерины растений, по-видимому, синтезируются из циклоартенола. К числу стеринов с дополнительным атомом углерода относится метиленциклоартенол — соединение, присутствующее в кожуре грейпфрута и во многих других растениях. Его боковая цепь имеет следующую структуру:
Другой стерин растении — кампестерин — содержит ∆5-ненасыщенное кольцо, как и холестерин, но боковая цепь имеет дополнительную метильную группу.
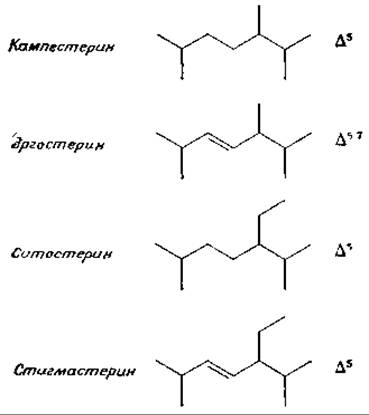
Эргостерин, характерный для грибов представитель стеринов, содержит ∆5,7 кольцевую систему, как и 7-дегидрохолестерин, а также дополнительную двойную связь в боковой цепи. Ситостерин и стигмастерин широко распространены среди высших растений. Отличительной особенностью этих стеринов служит присутствие дополнительной этильной группы в боковой цепи. Ситостерин образуется путем метилирования (при участии S-аденозилметионина) эргостерина. Стигмастерин является витамином для морских свинок, у которых он предотвращает окостенение суставов («фактор гибкости).