Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Типы реакций, катализируемых ферментами
Реакции множественного замещения и сопряжение реакций расщепления АТР с эндергоническими процессами
Общий механизм образования тиоэфиров, сложных эфиров и амидов
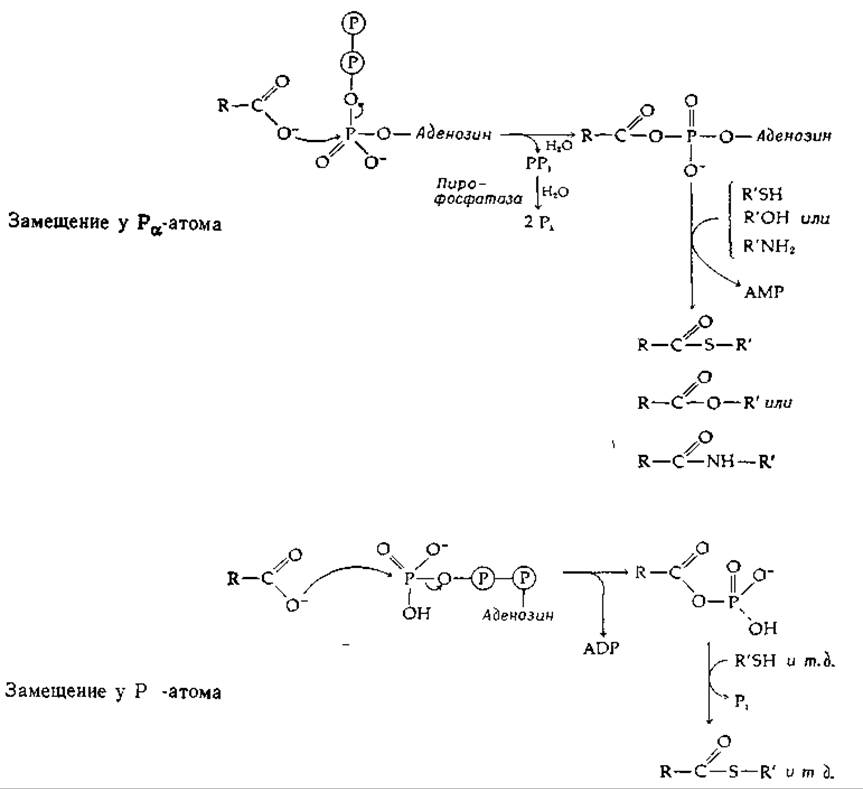
Последовательности реакций, показанные в уравнениях (7-29) и (7-30), представляют собой общий механизм, используемый клетками для присоединения карбоновых кислот к —ОН-,—SH- и—NН2-группам. Например, последовательность реакций (7-30) используется при образовании молекул аминоацил-тРНК, необходимых для синтеза белков. Механизм этих реакций показан в табл. 7-2. В зависимости от типа образующегося соединения (тиоэфир, сложный эфир или амид) реакции обозначены как S1A, S1B или S1C. Символы а и у указывают, в каком месте происходит расщепление АТР: при Ра или при Py. Например, образование ацетил-СоА у эукариотов протекает по механизму S1A(a). Понятно, что эта последовательность включает гидролиз неорганического пирофосфата (РРi) до неорганического фосфата (Pi), роль которого в сопряжении реакции расщепления АТР с биосинтезом рассмотрена ниже (гл. 11, разд. Б,2).
Несмотря на сделанные обобщения, каждый конкретный фермент имеет свои особенности. Действие сукцинил-СоА—синтетазы (сукцинат-тиокиназы) Е. coli, вероятно, осуществляется по механизму S1A(y), и в качестве промежуточного соединения выступает сукцинилфосфат:
![]()
Обозначение последовательностей реакций, использованное в настоящей книге:
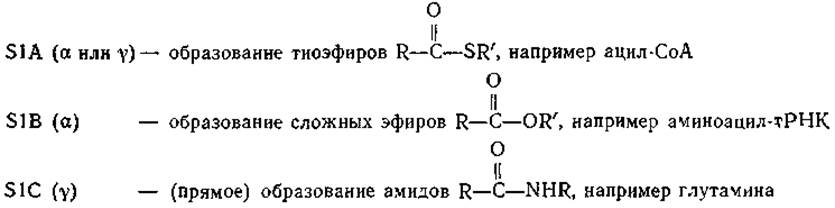
а3начение последовательных замещений у атома Р, а затем у атома С состоит в том, чтобы связывать молекулы небольших размеров в процессе, сопряженном с расщеплением АТР или родственного соединения. Расщепление АТР может происходить у Ра- или Ру-атома
Однако первой стадией может быть образование фосфофермента, в котором фосфатная группа присоединена к атому N3 боковой цепи остатка гистидина, принадлежащего а-субъединице тетрамера a2ß2[88. 88а, b].
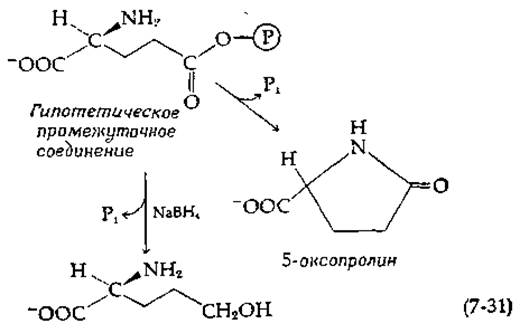
Синтез глутамина, катализируемый глутаминсинтетазой, вероятно, протекает по типу S1C(y). Однако прямых доказательств образования предполагаемого промежуточного продукта углутамилфосфата пока не имеется. Парциальные реакции изотопного обмена, которые в случае такого синтеза, по-видимому, должны были бы иметь место, не наблюдались [89]. Очевидно, образующийся в качестве промежуточного соединения ацилфосфат является короткоживущим соединением, и все три реагента должны одновременно присоединиться к ферменту прежде, чем активный центр станет способным к функционированию. О том, что образование ацилфосфата действительно происходит [90, 91], свидетельствует выделение внутреннего амида глутаминовой кислоты 5-оксопролина (пирролидонкарбоновой кислоты) и восстановление промежуточного продукта боргидридом натрия в спирт:
Поскольку прямое доказательство образования ацилфосфатов как промежуточных продуктов оказалось делом нелегким, было высказано предположение, что глутаминсинтетаза и некоторые другие ферменты, катализирующие реакции множественного замещения, могут обусловливать «полностью согласованную» реакцию. В этом случае обе стадии замещения должны протекать одновременно через единое «ключевое переходное состояние», как показано на приведенной ниже схеме для глутаминсинтетазы:
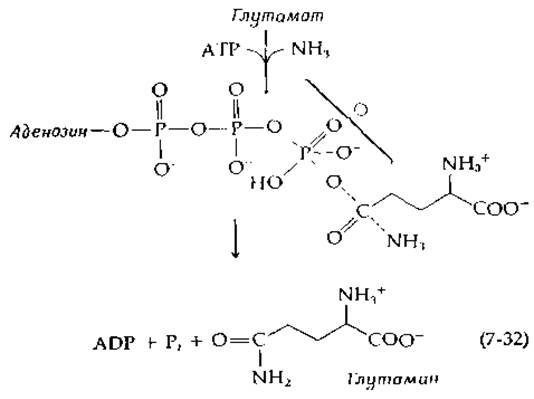
Однако до сих пор не получено ни одного сколько-нибудь убедительного довода в пользу существования такого механизма для какой-либо химической реакции. По-видимому, гораздо более вероятно, что ферментативные реакции протекают через ряд дискретных стадий.