Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Биохимическая генетика и синтез нуклеиновых кислот и белков
Хромосома эукариот и ее контроль
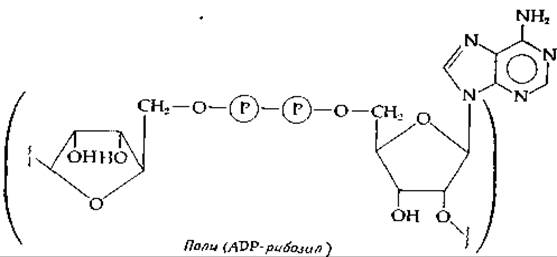
Поли (ADP-рибоза)
Необычный компонент клеточных ядер всех высших организмов образуется путем полимеризации ADP-рибозильных групп, образующихся из NAD+. Специальный фермент, катализирующий процесс полимеризации, вытесняет никотинамид и образует гликозидную связь между атомом С-1 рибозы, с которым был связан никотинамид, и атомом С-2' ADP-компонента следующей мономерной единицы. Значение поли(АDР-рибозильных групп) в настоящее время неясно, однако обнаружено, что они ковалентно связаны с ядерными белками, и также с цитоплазматическими компонентами [304, 305] (дополнение 15-Е).
Дополнение 15-Е
Токсичные белки; дифтерийный токсинa
До тех пор пока не удалось получить соответствующую вакцину, заражение Corynebacterium diphteriae вызывало одну из самых страшных детских болезней. Несмотря на то что возбудитель вызывал лишь образование поверхностных пленок в зеве, больные нередко умирали от тяжелого поражения многих органов. Причиной этого оказался активный термолабильный токсин белковой природы. Было обнаружено также, что бактерии продуцируют токсин лишь в том случае, если они заражены умеренным бактериофагомб, несущим ген tox, и при условии, что содержание неорганического железа в среде значительно снижено.
Дифтерийный токсин представляет собой белок с мол. весом ~62 000. Его минимальная летальная доза для морской свинки составляет всего лишь 0,16 мг/кг. Исследования, проведенные на культуре клеток, показали, что токсин блокирует включение аминокислот в белки в результате инактивации фактора элонгации EF-2, необходимого для транслокацин в рибосомах млекопитающих. Токсин действует аналогично ферменту, переносящему ADP-рибозильную группу от NAD+ к фактору EF-2:
![]()
Модифицированный фактор EF-2 реагирует с GTP обычным путем, однако образующийся при этом комплекс не способен принимать участие в транслокации. Для ускорения этой фатальной реакции достаточно, чтобы концентрация токсина в цитоплазме составляла всего лишь 10-8 М.
Каким же образом белковый токсин такого типа проникает в клетку? Имеются основания считать, что структура одного из участков белковой молекулы обладает способностью связываться с определенными участками клеточной, мембраны. Возможно, что связывание в этих участках стимулирует пиноцитоз. Токсин начинает действовать лишь после частичного протеолитического расщепления.
Каково происхождение гена tox и почему он переносится вирусом? Паппенхеймер и Джила высказали предположение, что этот ген каким-то образом образовался из гена эукариотической клетки, кодирующего функциональный белок. Этот ген внедрился в вирус и в ходе эволюции трансформировался в ген, детерминирующий синтез белкового токсина. Наличие в клеточном ядре поли (ADP-рибозы) (разд. И, 3) позволяет предложить одну из возможностей появления гена tox. NAD+ служит субстратом при синтезе этого ядерного полимера, а синтетаза катализирует разрыв рибозилникотинамидной связи с образованием новой гликозидной связи между 1-углеродом рибозы и 2-гидроксильной группой аденозина следующей мономерной единицы. Возможно, что именно ген синтетазы в результате модификации трансформировался в ген дифтерийного токсина.
О группе токсичных для бактерий белков (колицинов) уже шла речь в разд. Г, 7. Они, по-видимому, также связываются со специальными рецепторами на внешней мембране бактерий типа Е. coli. Нейландс и его сотрудники обнаружили, что у Е. coli рецептор колицина М служит также рецептором и для сидерохромного пептида — феррохрома (дополнение 14-В), и для бактериофага Т5. С этим же участком мембраны связывается антибиотик альбомицин. Существует предположение, что на ранних этапах эволюции у бактерий появились молекулы, обладающие способностью к образованию хелатных комплексов с железом, причем размер этих комплексов постепенно увеличился до такой степени, что они утратили способность диффундировать через наружную мембрану в клетку. В результате возникли специфические системы переноса, которые позднее были использованы фагами и штаммами, продуцирующими колицинв.
а Pappenheimer А. М., Jr., Gill D. М., Science, 182, 353—364 (1973).
б Eklund M. W., Poysky F. T., Reed S. M., Smith C. A., 172, 480—482 (1971).
в Lackey M., Wayne R., Neilands J. B., BBRC, 64, 687—693 (1975).
Вопросы и задачи
1. Как можно доказать, что передающиеся по наследству признаки действительно закодированы нуклеотидной последовательностью ДНК?
2. Опишите предложенную Уотсоном и Криком модель строения ДНК. Какие биологические и химические факты можно с ее помощью объяснить?
3. Приведите экспериментальные доказательства антипараллельной ориентации цепей в двухцепочечной ДНК. Охарактеризуйте специфичность ферментов, использованных для этого доказательства.
4. Если объединить все молекулы ДНК вашего организма в одну двухцепочечную спираль Уотсона — Крика, то какова будет ее: длина?
5. Опишите три типа РНК, участвующие в биосинтезе белков. Расскажите, что известно о химических и физических свойствах, нуклеотидном составе и о биосинтезе каждого из них?
6. Белок, являющийся репрессором lac-оперона, содержит только два трицтофановых остатка в положениях 190 и 209. При связывании с индуктором, изопропил-β-D-тиогалактозидом, происходит хотя и небольшое (на ~3 нм), но достоверное смещение максимума в спектре флуоресцепции белка в сторону длинных волн. Путем введения nonsense-мутаций и супрессии соответствующими супрессорными генами были получены модифицированные индукторы (см. разд. Г,5, Summer Н., Lu Р., Miller J. Н., JBC, 251, 3774—3779, 1976). При замене Тrр-190 на Туr смещение максимума в спектре флуоресценции при связывании индуктора оставалось таким же; если же Тrр-209 заменялся на Туr, то сдвига не наблюдалось. Прокомментируйте эти результаты. Можете ли вы предложить два возможных объяснения? Как можно определить, какое из них верно?
7. В бактериальной рибосоме диаметром 23 нм содержится 35% белка. Если предположить, что на долю белков приходится 35% объема, то сколько приблизительно белковых молекул со средним мол. весом 17 300 может находиться в рибосоме? (Считайте, что упаковка максимально компактна; см. также табл. 15-5.)
8. Молекулы транспортных РНК в процессе их синтеза и функционирования специфически взаимодействуют с несколькими ферментами. Напишите уравнения трех реакций, в процессе которых фермент узнает тРНК?
9. Назовите три функции определенных аминоацил-тРНК, не связанные с их участием в синтезе белка.
10. В глицинспецифической тРНК Е. coli содержится кодон GCC. Он может спариваться на рабосомах либо с кодоном GGU, либо с кодоном GGC. Как в настоящее время объясняют способность одной тРНК распознавать несколько разных кодонов?
11. Была выделена часть молекулы информационной РНК со следующей последовательностью оснований:
![]()
Как скажется на соответствующем пептиде, детерминированном этой информационной РНК, мутация, приводящая к замене в указанном стрелкой положении на С?
12. На 5'-конце многих молекул мРНК имеются длинные последовательности, не транслируемые рибосомной белоксинтезирующей системой. Опишите опыт, при помощи которого вы можете обнаружить начало информации в молекуле мРНК, считая, что располагаете чистым препаратом индивидуальной мРНК.
13. 7-метилгуанозин-5,-монофосфат ингибирует синтез белков в бесклеточной системе, выделенной из ретикулоцитов (Shafritz D. A. et al., Nature (London), 261, 291—294, 1976). На 5'-конце большинства молекул информационной РНК эукариотических клеток содержится 7-метилгуанозин. Однако удаление химическим путем этой группы из мРНК вируса везикулярного стоматита не предотвращает трансляции в бесклеточной системе из ретикулоцитов (Rose J. К., Lodish Н. F., 262, 32—37, 1976). Прокомментируйте значение этих наблюдений.
14. Дайте определение и опишите значение для биохимической генетики каждого из предложенных терминов:
|
Ауксотрофы по питательным веществам |
Комплементарность |
|
Метод реплик |
Супрессорные гены |
15. Сопоставьте следующие термины, используемые в биохимической генетике:
|
Трансформация |
Оперон |
|
Трансдукция |
Кодон |
|
Цистрон |
16. Опишите следующие формы ДНК (или ДНК-содержащих частиц):
|
Плазмида |
Фактор R |
|
Эписома |
Фактор F |
|
Колициногенный фактор |
Лизогенизирующий фаг |
17. Как может встраиваться в плазмиду кусок чужеродной ДНК? Почему включение ДНК эукариот в плазмиду имеет как биохимическое, так и этическое значение? Какую важную биохимическую проблему, возможно, удастся решить в будущем при помощи этого метода?
18. Как можно методом гетеродуплексного анализа определить локализацию в хромосоме включенной чужеродной ДНК?
19. а) В чем состоит различие между хромосомами Е coli и млекопитающих? Как можно обнаружить эти различия? б) Дайте определение эухроматина, гетерохроматина, сателлитной ДНК, повторов ДНК и гистонов. Укажите их возможные функции.
20. Дайте определение следующих типов мутантов:
|
С заменой пары оснований |
Температурочувствительные |
|
Со сдвигом рамки |
Nonsence |
21. Опишите химические принципы действия следующих мутагенных веществ или агентов:
|
Гидроксиламин |
Иодистый метил (слабый мутаген) |
|
5-бромоурацил |
Ультрафиолетовое излучение |
|
9-аминоакридин |
Бактериофаг |
22. Какие особые свойства вы можете предсказать для тирозиновой тРНК с антикодоном GψA, если в результате мутации она превратится в тРНК с антикодоном СψА?
23. После того как выделенная из обычной кукурузы митохондриальная ДНК была обработана рестриктирующей эндонуклеазой, а затем продукты гидролиза были подвергнуты электрофорезу в геле агарозы, на электрофореграмме было обнаружено около 50 полос. В электрофореграммах, полученных при аналогичном исследовании ДНК кукурузы с признаком мужской стерильности (разд. Г, 9,д), некоторые из полос, присутствовавших в электрофореграммах «нормальной» митохондриальной ДНК, обнаружить не удалось (Levings С. S., III, Pring D. R., Sciense, 193, 158—160, 1967). Прокомментируйте это наблюдение, а также тот факт, что при скрещивании нормальной кукурузы с кукурузой, несущей признак мужской стерильности, картина митохондриальной ДНК потомства всегда соответствует картине родителя женского пола.
24. Как бы вы объяснили тот факт, что у лошака более мягкий характер чем у мула?