Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Свет в биологии
Фотосинтез
Две фотосистемы
Различие между фотосинтезирующими бактериями и зелеными растениями стало еще более очевидным после экспериментов Р. Эмерсона и его сотрудников [79 b], выполненных в 1956 г. Было известно, что свет с длиной волны 650 нм намного более эффективен, чем свет с длиной волны 680 нм. Однако Эмерсон и др. показали, что сочетание света этих двух длин волн дает более высокую скорость фотосинтеза, чем свет с каждой из указанных длин волн по отдельности. Это позволило предположить, что существуют две разные фотосистемы. Фотосистема, известная теперь как фотосистема I, активируется далеким красным светом (~700 нм), тогда как фотосистема II — красным светом с более высокой энергией (~650 нм). Это положение подтверждается множеством разных фактов. Еще в 1937 г. Хилл [79с] показал, что фотосинтетическое образование О2 может идти с использованием мягких окислителей, таких, как феррицианид и бензохинон, а Г. Гаффрон [79d] обнаружил, что некоторые зеленые водоросли способны вести фотоокисление Н2 до протонов [уравнение (13-25)], используя электроны для восстановления NADP. Таким образом, фотосистема I может быть отделена от фотосистемы II.
Был найден мощный гербицид, дихлорофенилдиметилмочевина, блокирующий перенос электронов между этими двумя фотосистемами. В присутствии указанного соединения электроны могут поступать в фотосистему I от таких искусственных доноров, как аскорбиновая кислота или индофенольный краситель.
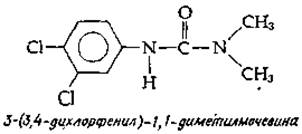
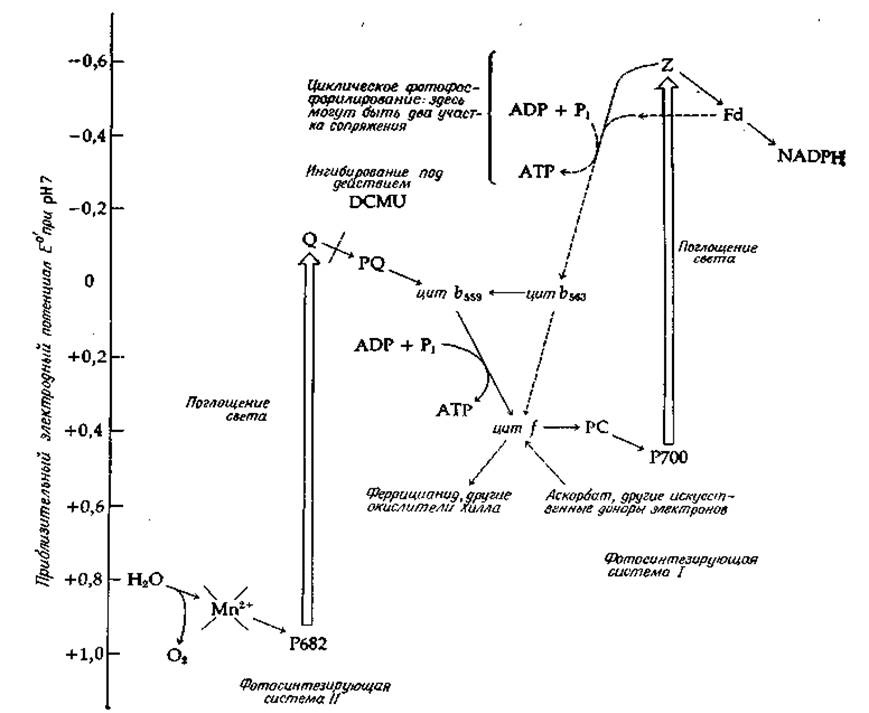
РИС. 13-18. Z-схема (от слова zigzag) фотовосстановления в хлоропластах; для перноса одного электрона необходимо затратить энергию двух квантов. Обозначения Р682 и Р700 — хлорофиллы реакционных центров; Q — тушитель флуоресценции Р681 и предполагаемый первый акцептор электронов; PQ — пластохинон; PC — пластоцианин; Z — первый акцептор электронов в фотосистеме I; Fd — ферредоксин; DCMU - дихлорофенилдиметилмочевина. Необходимо отметить, что положения Р682, Р700, O и Z на шкале значений Е0' точно не известны. Значения Е0' для Р682 и Р700 должны соответствовать паре (хлорофилл/хлорофильный катионный радикал) в окружений,) присущем их окружению в реакционных центрах; возможно, эти значения ниже, чем здесь указано. Неизвестны и окислительно-восстановительные потенциалы Q и Z; возможно, они выше, чем здесь указано.
В результате этих и других экспериментов был сформулирован ряд положений, послуживших основой для создания так называемой Z-схемы фотосинтеза (рис. 13-18). Перенос одного электрона через эту систему требует затраты энергии двух квантов света. Таким образом, для образования одной молекулы NADPH необходимо четыре кванта, а на каждый акт включения СО2 в состав углевода — восемь квантов. Большинство авторитетных специалистов в этой области считают, что для восстановления одной молекулы СO2 требуется по меньшей мере восемь-девять квантов.
Еще один важный эксперимент (поставленный Эмерсоном и Арнольдом [79 е]) был основан на использовании очень коротких вспышек света. При измерении квантового выхода фотосинтеза обнаружился поразительный факт: за один цикл работы фотосинтезирующего аппарата листьев на каждые 3000 молекул хлорофилла высвобождалась лишь одна молекула О2. Вместе с тем подсчеты показывали, что на каждую высвободившуюся молекулу О2 поглощалось лишь около восьми квантов света. Отсюда вытекало, что в поглощении одного кванта света участвует около 400 молекул хлорофилла, и, таким образом, можно было сделать вывод, что большое число молекул хлорофилла работает как единая светоулавливающая система, которая снабжает энергией один реакционный центр. Эта точка зрения в настоящее время получила всеобщее признание.