Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Биохимическая генетика и синтез нуклеиновых кислот и белков
Транскрипция молекул РНК
Транспортная РНК (тРНК)
Наиболее хорошо изученной формой РНК являются небольшие 4S-молекулы тРНК с мол. весом приблизительно 26000, состоящие из 75±5 нуклеотидов (рис. 2-24 и 15-8). Похоже, что по размерам и основным структурным характеристикам тРНК бактериальных и эукариотических клеток не различаются. Наличие «адаптеров», необходимых для переноса аминокислот в соответствующие участки матричной цепи мРНК, было предсказано еще до открытия тРНК. Предполагалось, что в молекуле «адаптора» должна иметься нуклеотидная последовательность, образующая антикодон, который должен точно располагаться против соответствующего кодона, в каком-то связывающем участке белоксинтезирующей системы. Как теперь известно, молекулы тРНК действительно обладают предсказанными свойствами, однако при изучении их химических свойств было обнаружено много неожиданного. Во-первых, молекулы тРНК оказались длиннее, чем это необходимо для формирования адаптеров, а во-вторых, многие основания, входящие в их состав, сильно модифицированы по сравнению с исходными соединениями [60].
Еще одна удивительная особенность, характерная для строения молекул тРНК, состоит в том, что в состав антикодонов входят не только «стандартные» основания. Так, например, в состав некоторых антикодонов входит гипоксантин (нуклеозидом которого является инозин).
На рис. 15-8, A приведено общепринятое изображение молекулы тРНК в виде «клеверного листа». Трехмерная структура тРНК, определенная независимо двумя группами исследователей [61, 62], схематически показана на рис. 15-8, Б.
Рассмотрим четыре «стебля», образованных нуклеотидными парами, связанными при помощи водородных мостиков (рис. 15-8). Один из этих «стеблей» заканчивается акцепторным концом, т. е. участком, в котором происходит связывание аминокислоты. Акцепторный конец присоединяет и переносит активированную аминокислоту, генерируемую в соответствии с уравнением (11-2). Три остальных «стебля» заканчиваются петлями, в состав которых обычно входит значительное число модифицированных оснований. Дигидроуридииовая петля, например, содержит 5,6-дигидроуридин в различном количестве и в различных положениях. В антикодоновой петле всегда находится антикодон, расположенный в клеверном листе на конце, прямо противоположном акцепторному концу. На стороне 5'-конца антикодона всегда находится U (на рисунке обведен кружком), за которым следует еще один пиримидиновый нуклеотид. Со стороны 3'-конца антикодона обычно находится так называемый «супермодифицированный» нуклеотид. ТφС-петля содержит специфическую нуклеотидную последовательность, от которой и происходит ее название. У бактерий последовательность ТφС была обнаружена во всех изученных бактериальных тРНК, участвующих в синтезе белка; в молекулах же инициаторной тРНК эукариотических клеток вместо этой последовательности может находиться последовательность UCG [63—65].
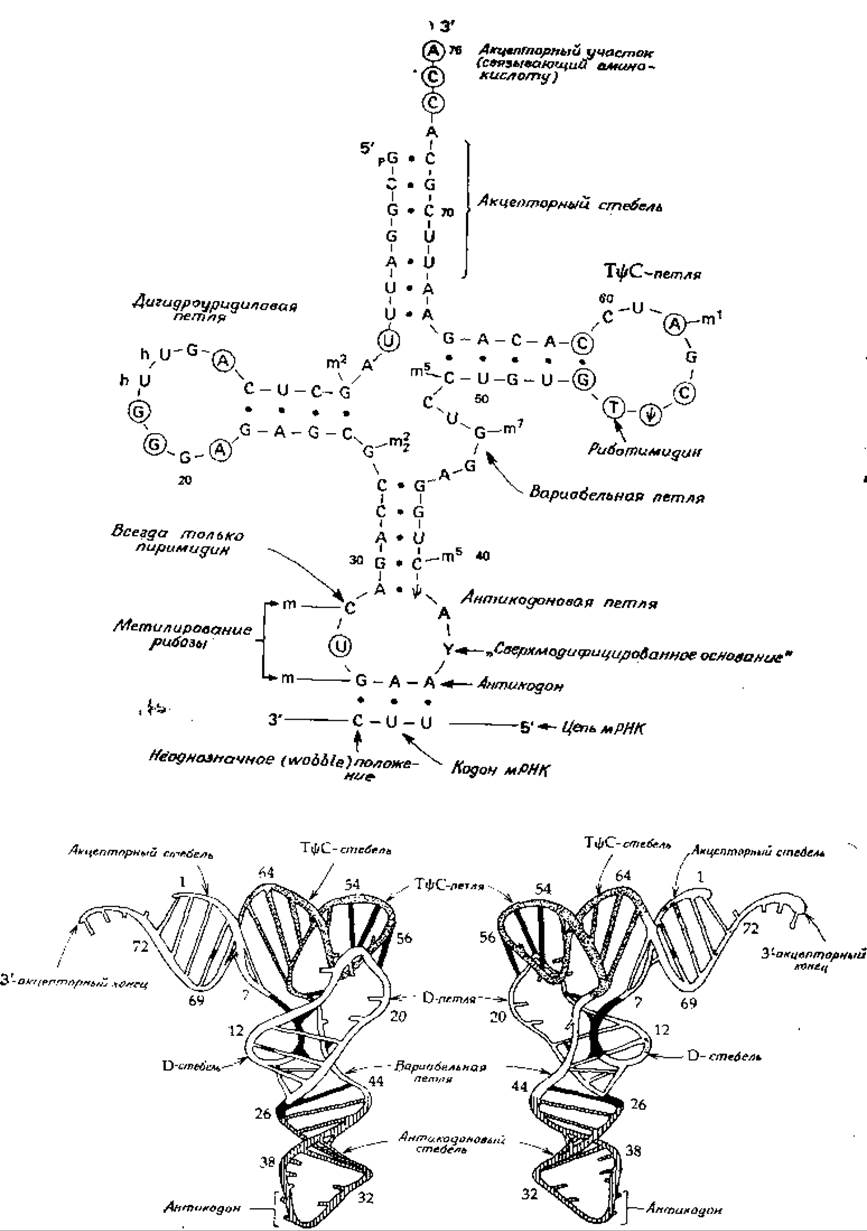
РИС. 15-8. Наверху. Схематическое изображение молекулы фенилаланиновой тРНК из дрожжей: структура клеверного листа. Точками обозначены водородные связи (две или три). Нуклеотиды, встречающиеся в молекулах почти всех тРНК, обведены кружком. Отмечены также и другие особенности, свойственные молекулам почти всех тРНК. В нижней части рисунка показано, как антикодон может взаимодействовать с кодоном мРНК Внизу. Трехмерная структура фенилаланиновой тРНК из дрожжей [61].
Гены, кодирующие синтез молекул тРНК, как у бактерий, так и у млекопитающих, объединены в кластеры, т. е. в определенные группы, при транскрипции которых образуются крупные молекулы — предшественники РНК, содержащие иногда молекулы более чем одного вида тРНК [66—69]. Для формирования зрелых молекул тРНК путем «разрезания и обстригання» («cutting and trimming») их предшественников необходимы по меньшей мере три различные нуклеазы [57, 66, 69].
Так, в случае тирозиновой тРНК1) Е. coli в молекуле непосредственного предшественника (рис. 15-9) содержится 129 нуклеотидов — на 44 больше, чем в молекуле зрелой тРНК. Из этих избыточных нуклеотидов 41 расположены с 5'-конца и три — с 3'-конца. Специальная нуклеаза (РНКаза Р) разрывает лишь одну связь в молекуле-предшественнике, отщепляя с 5'-конца фрагмент, состоящий из 41 нуклеотида. В результате действия другой нуклеазы (РНКаза PI1I) с противоположного конца отщепляется фрагмент, состоящий из трех нуклеотидов [66]. Дальнейшее разрушение отщепившихся фрагментов происходит под воздействием дополнительных ферментов [70].
Важным достижением химии явился синтез двухцепочечного фрагмента ДНК, кодирующего тирозиновую тРНК [71] Е. coli и ее предшественник [71а], осуществленный Кораной и его сотрудниками. Эти исследователи продолжили свою работу, чтобы охватить участок, терминирующий считывание гена, расположенный вне участка, кодирующего синтез CCA-конца тРНК. Была определена последовательность 23 нуклеотидов цепи ДНК, расположенных за участком, кодирующим синтез CCA-конца тРНК [72, 73] (рис. 15-9). Здесь следует отметить две особенности. Первая из них состоит в наличии участка, обладающего вращательной симметрией второго порядка (на рис. 15-9 он обозначен вертикальными линиями и точкой в центре), который может служить сигналом терминации. Вторая особенность связана с наличием двух участков, в которых короткие последовательности нуклеотидов повторяются, но в обратном направлении, например TGAAGT. Играют ли эти участки роль генетических сигналов, пока неизвестно2).
Участок из 29 нуклеотидов, расположенных перед геном тирозиновой тРНК, содержит оператор, последовательность которого также не установлена [75].
Процесс формирования окончательных молекул тРНК помимо «обстригання» предшественника нуклеазами сопровождается также значительными модификациями пуриновых и пиримидиновых оснований [64]. Известно шестьдесят или даже больше реакций, приводящих к таким модификациям; число и степень таких модификаций неодинаковы для разных видов. Строение некоторых модифицированных оснований показано на рис. 15-10. На примере уридина, приведенного на рис. 15-10, можно видеть, что возможны различные типы модификаций. Одной из наиболее типичных модификаций является метилирование, которое может происходить как по основанию, так и по 2'-гидроксильной группе сахарного остатка. Метилирование уридина по положению 5' приводит к образованию риботимидина. По этому же положению может быть модифицирован и цитидин. При восстановлении двойной связи между С-5 и С-6 образуется дигидроуридин. Замещение кислорода в положении 4 на серу дает 4-тиоуридин. На рис. 15-10 показано место возможного метилирования гуанозина. Для обозначения метилирования оснований нуклеиновых кислот часто используют символ m (диметилирование обозначают m2).
1) Минорная тирознновая тРНК, кодируемая супрессорным геном supF (разд. Г,6).
2) Для обозначения участков двухцепочечной ДНК, обладающих симметрией второго порядка, часто используют слово «палиндром» (гл. 2, разд. Г, 11). К сожалению, Корнберг [73] и другие исследователи воспользовались этим термином также и для обозначения примыкающих друг к другу участков одноцепочечной ДНК с обращенной последовательностью. Хотя такой палиндром должен как будто считываться одинаково в обоих направлениях, тем не менее ни одна ферментная система, движущаяся вдоль двухцeпочечной (или даже одноцепочечной) ДНК, не сможет, его прочитать oдинаково.
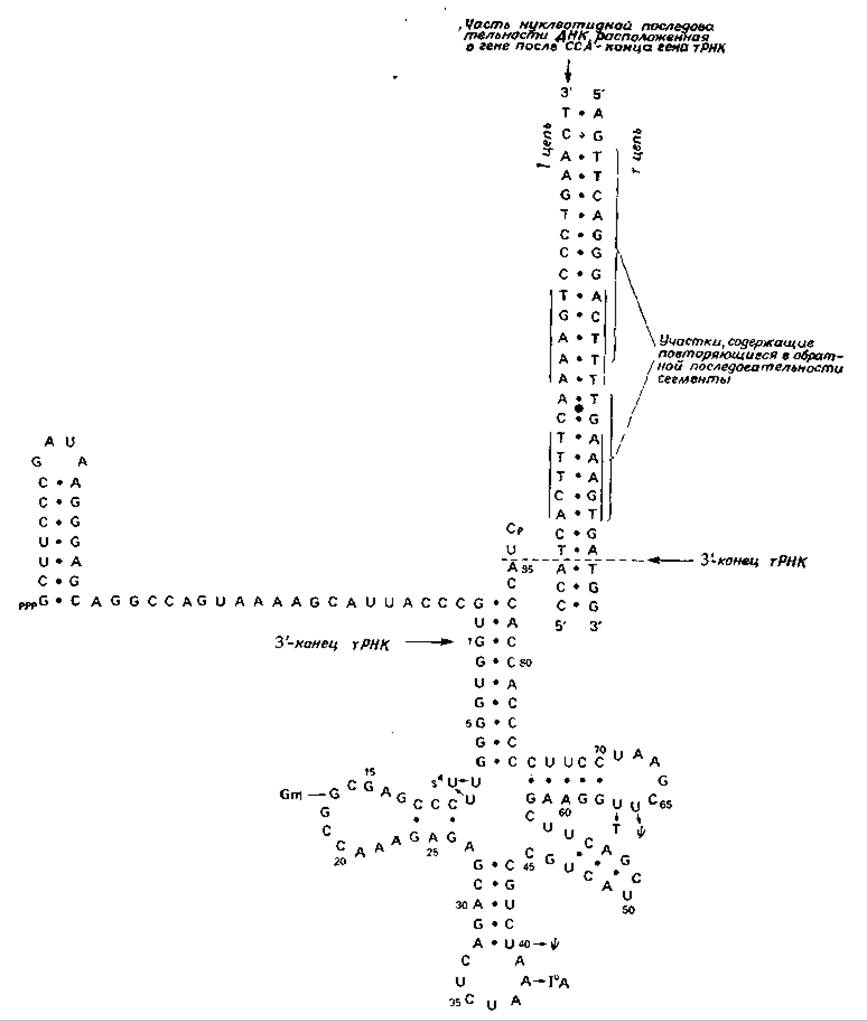
РИС. 15-9. Предполагаемая вторичная структура предшественника тирозиновой тРНК Е. coli. Нуклеотиды, которые обнаруживаются в зрелой тРНК в модифицированной форме, представлены на рисунке с соответствующими модификациями (Schaeffer К. Р-. Altman S., Sоll D., PNAS, 70, 3626—3630, 1973). Показана также часть нуклеотидной последовательности участка гена, кодирующего тРНК и расположенного после CCA-конца. Обратите внимание на участок, обладающий вращательной симметрией второго порядка (обозначен вертикальными линиями и точкой) и на два участка с обращенными нуклеотидными последовательностями.
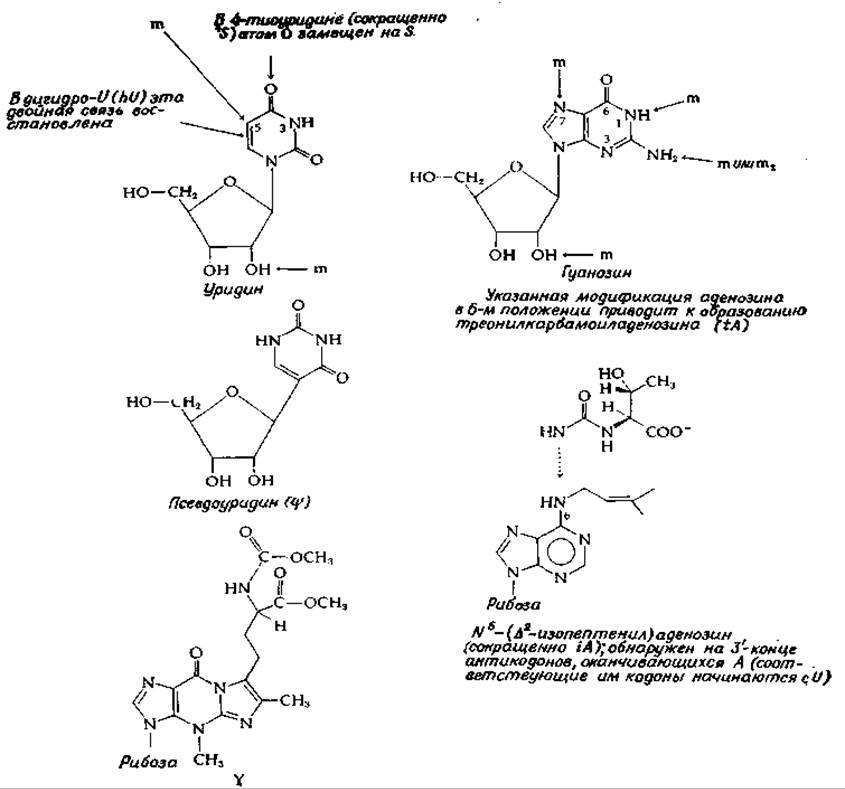
РИС. 15-10. Строение некоторых обнаруженных в молекулах тРНК нуклеозидов, содержащих модифицированные основания. Положения, в которых может осуществляться метилирование, обозначены буквой m.
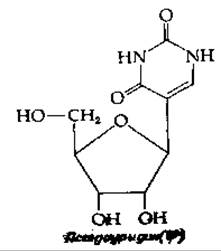
Большой интерес представляет собой превращение уридина в псевдоуридин (ψ). Известно, что псевдоуридин образуется в результате перегруппировки уридина, осуществляемой в первоначальном гранскрипте, однако химия такого превращения еще неясна. Существенным является то, что псевдоуридин, так же как и урацил, может «спариваться» с аденином. Основание, обозначаемое буквой Y, представляет собой сильно модифицированный гуанин. На рис. 15-10 показаны два «сверхмодифицированных» аденозина. На 3'-концах антикодонов, «спаривающихся» с кодонами, начинающимися с U, обнаруживается N6-изопептеннладенозин. Интересно, что у растений именно это соединение служит гормоном, известным под названием цитокинина (гл. 16, разд. А, 3).
Другой сильно модифицированный пурин — треонилкарбамонладенин — обычно примыкает к тому концу антикодонов, которые взаимодействуют с кодонами, начинающимися с А. Функция этих сильно модифицированных оснований неясна, однако похоже, что они нужны для правильного связывания тРНК с рибосомами. Специфический фермент, осуществляющий перенос изопентенильной группы с изопентилпирофосфата, был выделен в чистом виде (гл. 12, разд. 3) [76].
Интересная особенность метаболизма тРНК, которую мы до сих пор не рассматривали, связана с 3'-концевой группой, состоящей из трех нуклеотидов: ССА. Эта легко отщепляемая и вновь синтезируемая группа неизменно присутствует в молекулах всех тРНК. Скорость «оборота» этой группы достаточно высока для того, чтобы включить «в работу» приблизительно 20% молекул тРНК в промежуток между двумя делениями клетки, однако она значительно ниже той скорости, с которой молекулы тРНК участвуют в синтезе белка. Таким образом, этот процесс, по-видимому, непосредственно не связан с образованием пептидной связи.