Принципы структурной организации белков - Г. Шульц 1982
Способы свертывания и ассоциации полипептидных цепей
Агрегаты глобулярных белков
Контакты между субъединицами в дегидрогеназах
В 222-тетрамере необходимо образование только двух типов контактов из трех возможных. Рассмотрим типы контактов, которые могут возникать в симметричном олигомере, на примере четырех дегидрогеназ с известной структурой. s-Малат- и алкогольдегидрогеназы представляют собой димеры идентичных субъединиц с точечной группой 2, а лактат- и глицеральдегид-3-фосфатдегидрогеназы — тетрамеры идентичных субъединиц с точечной группой 222 (рис. 5.16, а). Димеры образуют контакт только одного типа вблизи поворотной оси 2-го порядка (рис. 5.18, а). В тетрамерах имеются три типа контактов, по одному на пересечении с каждой осью второго порядка (рис. 5.18, б). Отметим, что для стабилизации тетрамера необходимы только два из трех контактов.
Контакты 222-тетрамера можно представить тригональной диаграммой. Контакты тетрамера можно условно описать тетраэдром, показанным на рис. 5.18, б, а также двумерным графиком на рис. 5.18, в. Для идентификации субъединиц использован цветовой код, а поворотные оси второго порядка обозначены Р, Q и R. «Красная» субъединица считается базовой. Она образует контакты с «голубой», «желтой» и «зеленой» субъединицами, пересекающиеся соответственно осями Р, Q и R. Как видно из рис. 5.18, г, двумерный график удобно использовать для подсчета взаимодействий между остатками «красной» субъединицы с остатками других субъединиц.
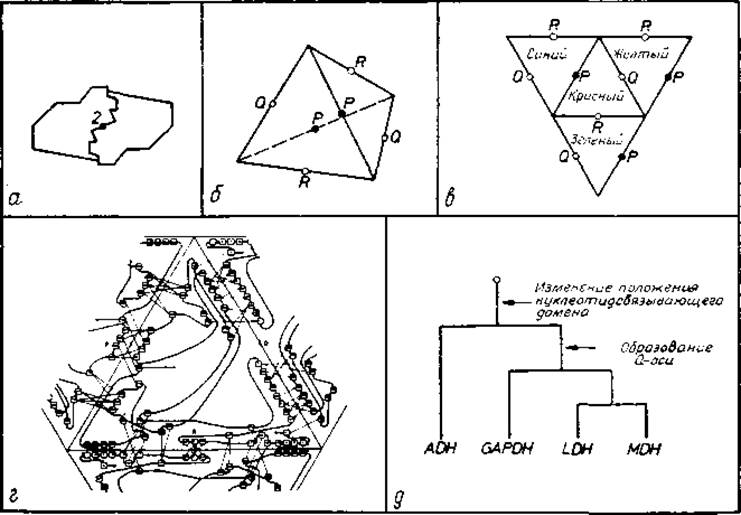
Рис. 5.18. Олигомерные ферменты — дегидрогеназы. а — схема симметричного димера, например алкогольдегидрогеназы; имеется поверхность раздела только одного типа. Контактирующие поверхности комплементарны (система ключ—замок [44]). б — тетрамер симметрии 222 представлен тетраэдром. Указаны три поворотные оси 2-го порядка: Р, Q и R. Каждая субъединица представлена треугольником, а каждый контакт между субъединицами — ребром, в — плоское представление тетраэдра и контактов между субъединицами, г — использование плоского представления для идентификации атомных контактов между субъединицами лактатдегидрогеназы [264]. д — дерево эволюции дегидрогеназ, построенное на основе анализа контактов между субъединицами [264]. ADH — алкогольдегидрогеназа, GAFDH — глицеральдегид-3-фосфатдегидрогеназа, LDH — лактатдегидрогеназа, MDH — s-малатдегидрогеназа.
Контакты выявляют эволюционную взаимосвязь. В работах Россмана и сотр. [264, 265] было показано, что четыре дегидрогеназы эволюционно связаны и что сравнение контактов между субъединицами позволяет выявить некоторые детали этой взаимосвязи. Эти контакты можно сопоставить, поскольку все четыре дегидрогеназы имеют одинаковый нуклеотидсвязывающий домен (рис. 5.17, б), составляющий приблизительно половину субъединицы. Такой домен можно использовать в качестве базового. Для лактат- и s-малатдегидрогеназы характерно почти одинаковое свертывание цепей, что свидетельствует о родственности этих белков. Глицеральдегид-3-фосфат, лактат- и s-малатдегидрогеназы образуют Q-осевой контакт (показано на рис. 5.18, г). Соответственно димер «красное» — «желтое» должен был хорошо сохраниться в процессе эволюции белка. Что касается Р- и R-осевых контактов дегидрогеназ, то они не проявляют никакого сходства. Кроме того, нет подобия между контактом в алгокольдегидрогеназе и каким-либо другим контактом*. Построенная на основании этих данных схема эволюционной взаимосвязи приведена на рис. 5.18, д.