БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 16. ГЛИКОГЕН И ОБМЕН ДИСАХАРИДОВ
16.8. Ветвящий фермент образует альфа-1,6-связн
Гликоген-синтаза катализирует только синтез α-1,4-связей. Для образования α-1,6- связей, благодаря которым гликоген становится разветвленным полимером, требуется другой фермент. Ветвление имеет важное значение в том отношении, что оно повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликоген-фосфорилазы и гликоген-синтазы, Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена.
Ветвление происходит после присоединения некоторого количества глюкозильных остатков через α-1,4-связь под действием гликоген-синтазы. Ветвь создается путем разрушения α-1,4-звена и образования α-1,6-звена, что представляет собой реакцию, отличную от реакции, устраняющей ветвление. Блок остатков, обычно в количестве семи, переносится ближе к внутренней части молекулы. Ветвящий фермент, катализирующий эту реакцию, обладает очень строгой специфичностью. Блок, состоящий из семи (или около этого) остатков, должен включать в себя невосстанавливающий конец и происходить из цепи длиной минимум в одиннадцать остатков. Кроме того, новая точка ветвления должна быть отдалена от предсуществующей точки ветвления по меньшей мере на четыре остатка.
16.9. Гликоген-очень эффективная резервная форма глюкозы
Какова цена превращения глюкозо-6-фосфата в гликоген и обратного превращения гликогена в глюкозо-6-фосфат? Соответствующие реакции уже были описаны, за исключением приведенной ниже реакции 5, представляющей собою регенерацию UTP. UDP фосфорилируется за счет АТР в реакции, катализируемой нуклеозид-дифосфокиназой.
1) Глюкозо-6-фосфат → Глюкозо-1-фосфат
2) Глюкозо-1-фосфат + UTP → UDP-глюкоза + РРi
3) РРi + Н2O → 2Рi
4) UDP-глюкоза + Гликоген n → Гликоген n+1 + UDP
5) UDP + ATP → UTP + ADP
Суммарная реакция: Глюкозо-6-фосфат + АТР + Гликоген n + Н2O → Гликоген n+1 + ADP + 2Рi
Таким образом, одна высокоэнергегическая фосфатная связь расходуется при включении глюкозо-6-фосфата в гликоген. Выход энергии при распаде гликогена чрезвычайно высок. Около 90% остатков подвергаются фосфоролитическому расщеплению с образованием глюкозо-1-фосфата, который превращается в глюкозо-6- фосфат без энергетических затрат. Остальные 10% остатков принадлежат ветвям и расщепляются гидролитически. Одна молекула АТР используется для фосфорилирования каждой из этих молекул глюкозы в глюкозо-6-фосфат. Полное окисление глюкозо-6-фосфата дает тридцать семь молекул АТР, а при запасании потребляется чуть больше одной молекулы АТР на молекулу глюкозо-6-фосфата, так что общая эффективность запасания составляет около 97%,
16.10. Циклический АМР занимает центральное положение в координированном контроле синтеза и распада гликогена
Существование раздельных путей для синтеза и распада гликогена означает, что эти процессы должны подчиняться строгой регуляции. Полное проявление активности обеих групп реакций в одно и то же время привело бы к «непроизводительному» гидролизу АТР. В действительности синтез и расщепление гликогена координированно регулируются таким образом, что гликоген-синтаза оказывается почти неактивной при полной активности фосфорилазы и наоборот. На обмен гликогена большое влияние оказывают специфические гормоны. Полипептидный гормон инсулин (разд. 2.6) повышает способность печени синтезировать гликоген. Механизм действия инсулина пока не раскрыт. Высокое содержание инсулина в крови говорит о состоянии сытости, тогда как его низкое содержание является сигналом голода (разд. 23.6). Значительно лучше изучен механизм действия адреналина и глюкагона, эффект которых противоположен эффекту инсулина. Мышечная активность или подготовка к ней приводит к высвобождению адреналина мозговым веществом надпочечников. Адреналин выраженно стимулирует распад гликогена в мышцах и в меньшей степени-в печени. Печень более чувствительна к глюкагону, полипептидному гормону, секретируемому α-клетками поджелудочной железы при низком содержании сахара в крови. Глюкагон повышает содержание сахара в крови, стимулируя распад гликогена в печени.
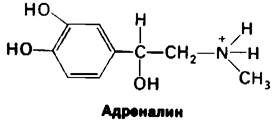

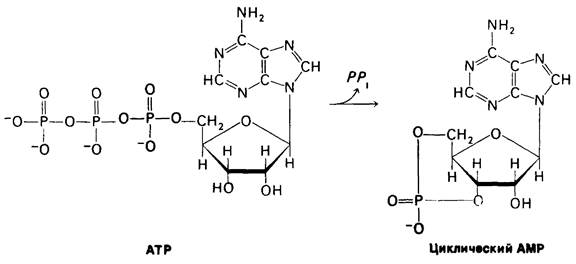
Эрл Сазерланд (Earl Sutherland) установил, что действие адреналина и глюкагона на метаболизм опосредуется циклическим АМР. Это открытие привело к представлению об универсальном присутствии циклического АМР во всех формах жизни и о его ключевой роли в регуляции биологических процессов (гл. 35). Синтез этой регуляторной молекулы из АТР катализируется аденилатциклазой, ферментом, связанным с плазматическими мембранами. Процесс ускоряется последующим гидролизом пирофосфата.
Адреналин и глюкагон не проникают в свои клетки-мишени. Они связываются с плазматическими мембранами и стимулируют аденилатциклазу (разд. 35.3). Повышенное внутриклеточное содержание циклического АМР запускает ряд реакций, приводящих к активации фосфорилазы и ингибированию гликоген-синтазы. Мы рассмотрим теперь структурную основу регуляции активности указанных ключевых ферментов обмена гликогена и затем перейдем к каскаду реакций, устанавливающих связь между этими ферментами и циклическим АМР.