БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 24 ДНК: ГЕНЕТИЧЕСКАЯ РОЛЬ, СТРУКТУРА И РЕПЛИКАЦИЯ
24.2. Трансформация пневмококков с помощью ДНК показала, что гены состоят из ДНК
Бактерии пневмококки сыграли важную роль в открытии генетической функции ДНК.
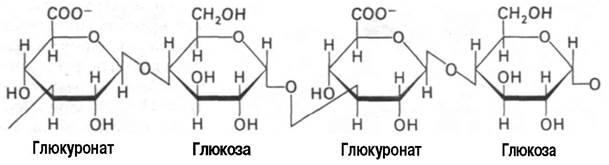
Пневмококки обычно окружены слизистой блестящей полисахаридной капсулой. Этот наружный слой имеет существенное значение для проявления патогенности
бактерий. Они вызывают пневмонию у человека и других чувствительных млекопитающих. Мутанты, лишенные полисахаридной оболочки, не патогенны. Бактерии дикого типа обозначают буквой S (от англ. smooth - гладкий), так как они образуют гладкие колонии, а мутантные бактерии, не имеющие капсулы, буквой R (от англ. rough - шероховатый), так как они образуют шероховатые колонии. У одной группы R-мутантов отсутствует фермент дегидрогеназа, превращающий UDР-глюкозу в UDР-глюкуронат. Фермент необходим для синтеза капсульного полисахарида, который у этих пневмококков состоит из чередующейся последовательности остатков глюкозы и глюкуроната:
В 1928 г. Фред Гриффит (Fred Griffith) обнаружил, что непатогенный R-мутант можно трансформировать в патогенную S-форму следующим путем. Он инъецировал мышам смесь живых бактерий R- формы и убитых нагреванием пневмококков S. Поразительное открытие Гриффита состояло в том, что эта смесь вызывала гибель мышей, хотя ни живые R-пневмококки, ни убитые нагреванием S-пневмококки мышей не убивали. Кровь погибших мышей содержала живые S-пневмококки. Следовательно, убитые нагреванием 8- пневмококки каким-то образом трансформировали живые R-пневмококки в живые S-пневмококки. Это изменение было стабильным: трансформированные пневмококки давали патогенное потомство S-формы. Затем было установлено, что такая трансформация R → S может происходить in vitro. Некоторые клетки в растущей культуре R-формы трансформировались в S-форму при добавлении бесклеточного экстракта убитых нагреванием пневмококков S. Это открытие заложило основу для
Рис. 24.1. Схема структуры ДНК. Сахарофосфатный остов показан черным цветом, а пуриновые и пиримидиновые основания разноцветные изучения химической природы трансформирующего начала
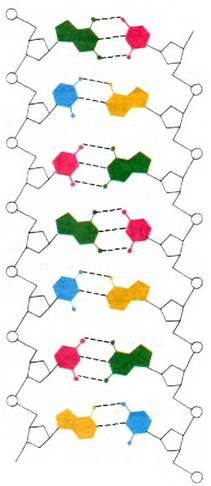
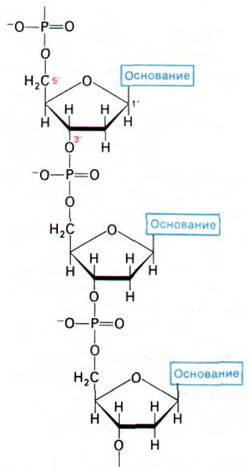
Рис. 24.2. Строение одной цепи ДНК. Показана часть цепи
Исследователи стали фракционировать бесклеточный экстракт убитых нагреванием пневмококков S и определять трансформирующую активность его компонентов (рис. 24.3). В 1944 г. Освальд Эйвери, Колин Мак-Леод и Маклин Мак-Карти (Oswald Avery, Colin MacLeod, Maclyn McCarty) опубликовали результаты своей работы. Оказалось, что «нуклеиновая кислота дезоксирибозного типа - основное действующее начало трансформирующего экстракта пневмококка типа III.» Экспериментальные доказательства этого заключения состояли в следующем: 1) при элементном химическом анализе очищенного, высокоактивного трансформирующего на-
чала результат был близок к расчетному для ДНК; 2) оптические, электрофоретические свойства, поведение при ультрацентрифугировании и диффузия очищенного материала соответствовали свойствам ДНК; 3) при экстрагировании белков или липидов не происходило потери трансформирующей активности; 4) трипсин и химо- трипсин не влияли на трансформирующую активность; 5) рибонуклеаза (которая, как известно, гидролизует рибонуклеиновую кислоту) не влияла на трансформирующее начало; 6) при добавлении дезоксирибонуклеазы трансформирующая активность, наоборот, терялась.
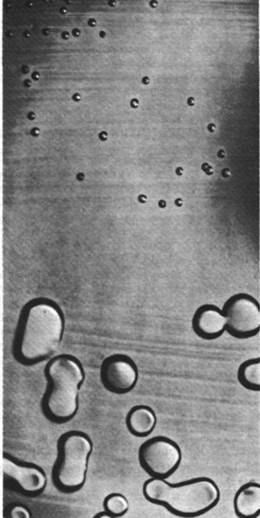
Рис. 24.3. Трансформация непатогенного пневмококка R (мелкие колонии) и возникновение патогенного пневмококка S (крупные блестящие колонии) под действием ДНК из убитых нагреванием пневмококков S
Эта работа - памятная веха в истории биохимии. До 1944 г. считалось, что генетическую информацию несут хромосомные белки, а ДНК играет вторичную роль. Такая преобладающая точка зрения была решительно опровергнута открытием строго доказанного факта, что очищенная ДНК обладает генетической специфичностью. В 1943 г. Эйвери (Avery) живым языком описал это исследование и его последствия в письме, направленном брату в другой университет (рис. 24.4).
Рис. 24.4. Из письма Освальда Эйвери брату Рою, написанного в мае 1943 г

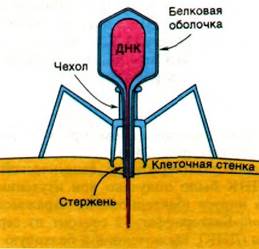
Новое подтверждение генетической роли ДНК было получено при изучении одного вируса (бактериофага), заражающего Е. coli. Бактериофаг Т2 состоит из сердцевины (ДНК), заключенной в белковую оболочку. В 1951 г. Роджер Херриот (Roger Herriott) предположил, что «вирус, очевидно, действует, как крошечный шприц для подкожных инъекций, наполненный трансформирующим началом; вирус как таковой никогда не проникает в клетку; только отросток вступает в контакт с клеткой-хозяином и, возможно, ферментативно проделывает небольшое отверстие в наружной мембране (рис. 24.5). Затем нуклеиновая кислота из головки вируса перетекает внутрь клетки». Чтобы проверить это предположение Альфред Херши и Марта Чейз (Alfred Hershey, Martha Chase) поставили следующий опыт. Фаговую ДНК пометили радиоактивным изотопом 32Р, а белковую оболочку - изотопом 35S. Эти метки весьма специфичны, так как ДНК не содержит серы, а в белковой оболочке нет фосфора. Культуру Е. coli заразили помеченным фагом, который за непродолжительное время инкубации прикреплялся к бактерии. Суспензию обрабатывали в течение нескольких минут в гомогенизаторе Уоринга при 10000 об/мин. В этих условиях зараженные фагом клетки подвергались воздействию значительных сил сдвига, которые разрушали связи между вирусами и бактериями. Затем суспензию центрифугировали, чтобы осадить бактерии на дно пробирки. Полученный осадок содержал зараженные бактерии, а надосадочная фракция - более мелкие частицы. Исследуя содержание 32Р и 35S в осадке и надосадочной фракции, определяли локализацию фаговой ДНК и белка оболочки. В результате были получены следующие данные.
Рис. 24.5. Схема бактериофага Т2, вводящего свою ДНК в бактериальную клетку
1. Большая часть фаговой ДНК обнаруживалась в бактериях.
2. Большая часть фагового белка обнаруживалась в надосадочной фракции.
3. Обработка в гомогенизаторе почти не влияет на способность зараженных бактерий продуцировать вирусное потомство.
Дополнительные эксперименты показали, что менее 1% 35S было перенесено из родительских фаговых частиц фаговому потомству; 30% родительской 32Р-метки, наоборот, обнаруживалось в потомстве. Эти простые, убедительные эксперименты показали, что фаг Т2 можно физически разделить на генетическую и негенетическую части... Серусодержащий белок покоящихся фаговых частиц содержится только в защитной оболочке, которая обеспечивает прикрепление фаговой частицы к бактериальной клетке и функционирует в качестве приспособления для введения фаговой ДНК в клетку. Этот белок, возможно, не несет никакой функции, необходимой для роста фаговых частиц внутри клетки. ДНК же выполняет какую-то функцию в размножении фага. Представленные эксперименты не позволяют сделать более далеко идущие выводы относительно химической природы наблюдаемых явлений».
Осторожный тон этого вывода не должен умалять его значения. Вскоре генетическая роль ДНК стала общепризнанной. Эксперименты Херши и Чейз убедительно подтвердили факты, открытые восемью годами раньше Эйвери, Мак-Леодом и МакКарти на другой системе. Дополнительные доказательства были получены при исследовании содержания ДНК в отдельных клетках. Они показали, что для данного вида содержание ДНК одинаково во всех
клетках с диплоидным набором хромосом. Гаплоидные клетки имеют вполовину меньше ДНК.