БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 24 ДНК: ГЕНЕТИЧЕСКАЯ РОЛЬ, СТРУКТУРА И РЕПЛИКАЦИЯ
24.3. Гены некоторых вирусов состоят из РНК
Гены всех прокариотических и эукариотических организмов построены из ДНК, гены вирусов - из ДНК или из РНК. Вирус табачной мозаики, заражающий листья
растений табака, - один из наиболее изученных РНК-содержащих вирусов. Он образован одной молекулой РНК, окруженной белковой оболочкой из 2130 одинаковых субъединиц (обсуждение структуры и сборки вирусов см. в гл. 30). Обработав вирус фенолом, можно отделить белок от РНК. Изолированная вирусная РНК инфекционна, тогда как изолированный белок лишен инфекционности. Использование синтетических гибридных вирусных частиц еще раз показало, что генетическая специфичность вируса заключается только в его РНК. Существует множество штаммов вируса табачной мозаики. Был получен синтетический гибридный вирус из РНК штамма 1 и белка штамма 2. Другой такой вирус был приготовлен из РНК штамма 2 и белка штамма 1. После заражения клетки гибридным вирусом вирусное потомство всегда состояло из РНК и белка, соответствующих специфичности РНК гибридного вируса, использованного для заражения.
24.4. Двойная спираль ДНК Уотсона-Крика
В 1953 г. Джеймс Уотсон и Френсис Крик (James Watson, Francis Crick) установили трехмерную структуру ДНК и сразу же предложили возможный механизм ее репликации. Это блестящее достижение стоит в ряду важнейших событий в истории биологии, так как оно открыло путь к пониманию функции гена на молекулярном уровне. Уотсон и Крик проанализировали картины дифракции рентгеновских лучей на нити ДНК (рис. 24.6), полученные Розалиндой Франклин и Морисом Уилкинсом (Rosalind Franklin, Maurice Wilkins), и предложили структурную модель, которая в основных своих чертах оказалась верной. Характерные особенности этой модели сводятся к следующему.
1. Две спиральные полинуклеотидные цепи закручены вокруг общей оси. Цепи направлены в противоположные стороны (рис. 24.7).
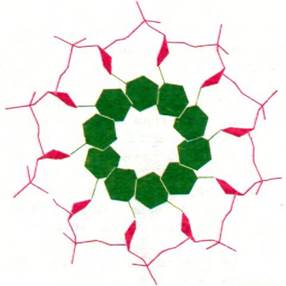
2. Пуриновые и пиримидиновые основания расположены внутри спирали, а остатки фосфата и дезоксирибозы - снаружи (рис. 24.8). Плоскости оснований перпендикулярны оси спирали. Плоскости остатков сахара расположены почти под прямым углом к основаниям.
3. Диаметр спирали 20 А. Расстояние
между соседними основаниями вдоль оси спирали 3,4 А, они повернуты относительно друг друга на 36°. Таким образом, на один виток спирали каждой из цепей приходится 10 нуклеотидов, что соответствует 34 А.
4. Две цепи удерживаются вместе водородными связями между парами оснований. Аденин всегда спаривается с тимином, гуанин - с цитозином (рис. 24.9 и 24.10).
5. На последовательность оснований в полинуклеотидной цепи не накладывается никаких ограничений. Определенная последовательность оснований несет конкретную генетическую информацию.
Рис. 24.6. Фотография дифракции рентгеновских лучей на влажной нити ДНК (В-форма). Крест в середине картины характерен для спиральной структуры. Интенсивные меридианальные рефлексы возникают в результате дифракции на стопке пар оснований, расположенных на расстоянии 3,4 А друг от друга
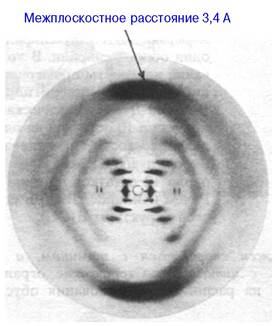
Рис. 24.7. Схема модели двухспиральной ДНК. Вся структура повторяется через 34 А, что соответствует 10 остаткам в каждой цепи
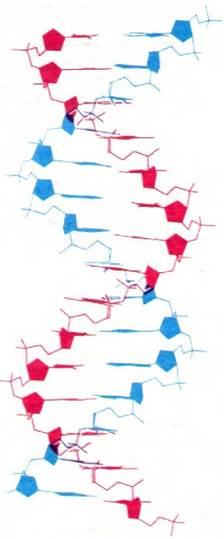
Рис. 24.8. Схема одной из цепей двойной спирали ДНК (вид по оси спирали). Основания - в данном случае только пиримидины - расположены внутри, а сахарофосфатный остов - снаружи. Отчетливо видно, что структура имеет ось симметрии десятого порядка. Основания обозначены зеленым цветом, сахара - красным
Важнейшее свойство двойной спирали - специфичность спаривания оснований. Исходя из стерических ограничений и способности к образованию водородных связей, Уотсон и Крик постулировали, что аденин должен спариваться с тимином, а гуанин - с цитозином. Стерические ограничения на расположение основания обусловлены периодическим строением спиральной структуры сахарофосфатного остова обеих полинуклеотидных цепей. Гликозидные связи, которыми присоединяется к остову пара оснований, отстоят друг от друга на расстояние 10,85 А (рис. 24.11). Пара пиримидин—пурин прекрасно укладывается в это пространство. В то же время для двух пуринов этого расстояния недостаточно. Для двух пиримидинов места более чем достаточно, но они оказались бы слишком далеко друг от друга, чтобы образовать водородные связи. Следовательно, одно из оснований в паре в двойной спирали ДНК всегда должно быть пурином, а другое - пиримидином. Кроме того, на спаривание оснований накладывают ограничения правила образования водородных связей. Атомы водорода в пуриновых и пиримидиновых основаниях занимают вполне определенные положения. Аденин не может спариваться с цитозином, так как тогда на месте одной связи оказалось бы два атома водорода, а на месте другой - ни одного. Точно так же гуанин не может связываться с тимином. Аденин, наоборот, образует с тимином две водородные связи, а гуанин - три водородные связи с цитозином (рис. 24.9 и 24.10). Ориентация этих водородных связей и расстояние между ними оптимальны для сильного взаимодействия между основаниями.
Рис. 24.9. Модель пары оснований аденин—тимин
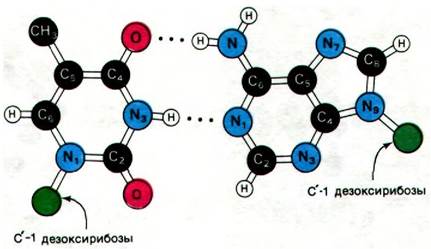
Рис. 24.10. Модель пары основании гуанин—цитозин
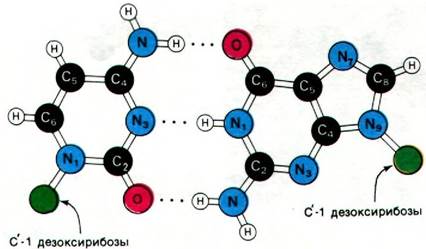
Рис. 24.11. Наложение АТ-пары оснований (обозначена желтым цветом) на GС-пару (обозначена синим цветом). Обратите внимание, что положения гликозидных связей и С'-1 атома дезоксирибозы (обозначен зеленым) в двух типах пар оснований почти одинаковы
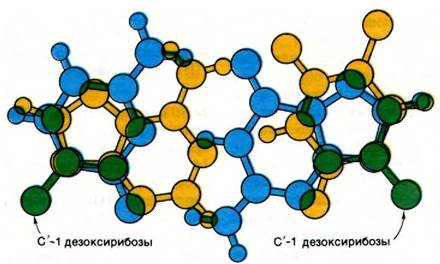
Формы ДНК —
А-форма ДНК: двухспиральная ДНК, содержащая примерно 11 остатков на один оборот спирали. В этой правозакрученной спирали плоскости пар оснований повернуты примерно на 20° относительно перпендикуляра к оси спирали. Образуется при дегидратации В-формы ДНК.
В-форма ДНК: классическая уотсон-криковская двойная спираль, содержащая примерно 10 остатков на один оборот спирали. В этой правозакрученной спирали плоскости пар оснований перпендикулярны оси спирали.
Z-форма ДНК: левозакрученная двухспиральная форма ДНК, содержащая около 12 остатков на один оборот. Эта структура предложена Александром Ричем (Alexander Rich) и его сотрудниками на основе изучения кристаллической структуры d(CG)3.
Эта схема спаривания оснований получила убедительное подтверждение в результате проведенных ранее исследований нуклеотидного состава ДНК разных видов. В 1950 гг. Эрвин Чаргафф (Erwin Chargaff) обнаружил, что соотношение аденина с тимином и гуанина с цитозином у всех исследованных видов было близко к 1. Значение этого факта оставалось неясным, пока не была предложена модель Уотсона-Крика (рис. 24.12 и 24.13). Только тогда стало понятным, что эта закономерность отражает важнейшее свойство структуры и функции ДНК - специфичность спаривания оснований.
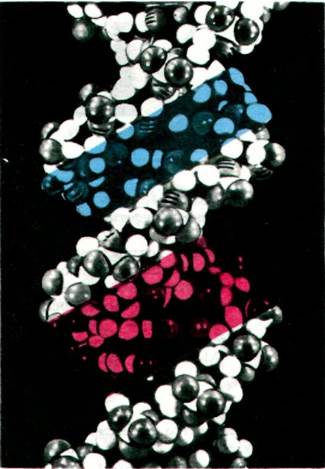
Рис. 24.12. Модель двухспиральной молекулы ДНК. Показаны три пары оснований. Обратите внимание, что две цепи ориентированы в противоположном направлении
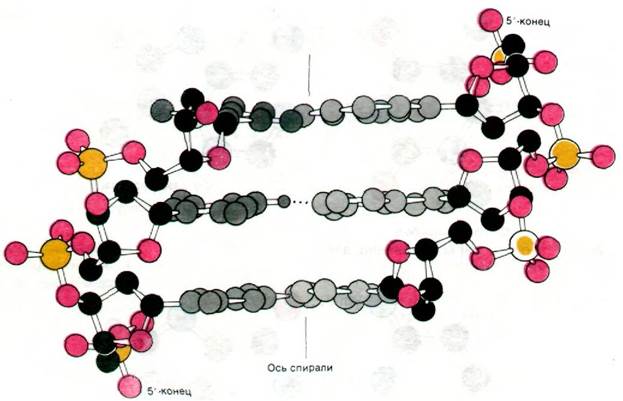
Рис. 24.13. Пространственная модель двойной спирали ДНК. Отчетливо видны большая бороздка (показана красным цветом) и малая бороздка (показана синим цветом). Обратите внимание, что часть каждого основания доступна для взаимодействия с другими молекулами