БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 29. ЭУКАРИОТИЧЕСКИЕ ХРОМОСОМЫ И ВЫРАЖЕНИЕ ГЕНОВ У ЭУКАРИОТ
29.20. Три вида рибосомных РНК образуются в результате процессинга одного первичного транскрипта
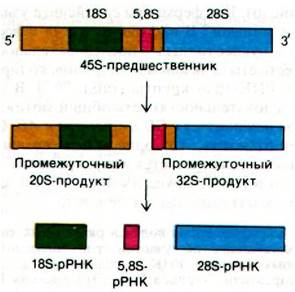
Молекулы РНК, синтезируемые тремя РНК-полимеразами, называются первичными транскриптами. Эти новообразованные РНК в очень большой степени модифицируются в ядре, прежде чем они переходят в цитозоль в виде зрелых молекул рРНК, тРНК и мРНК. Образование рибосомных РНК из первичного транскрипта было изучено детально, лучше, чем процессинг каких-либо других видов РНК. Выше уже обсуждалось, что гены 18S-, 5,8S- и 28S-pPHK сгруппированы в единый кластер и этот кластер генов повторяется тандемно много раз. В ядрышках эта группа из трех генов транскрибируется РНК-полимеразой I и дает 45S-PHK (рис. 29.30).
Рис. 29.30. Если расправить молекулу 45S- предшественника рибосомной РНК из клеток HeLa и исследовать ее с помощью электронного микроскопа, видна весьма характерная картина шпилек и петель. Благодаря этому можно картировать расположение молекул 28S- и 18S-PHK, образующихся из этого предшественника. А - электронная микрофотография 45S-предшественника. Б - схематическое изображение молекулы, изображенной на рис. А

Предшественник длиной 13 kb подвергается ферментативной модификации и расщеплению и дает зрелые 18S-, 5,8S- и 28S-pPHK.
(рис. 29.31). Примерно 100 нуклеотидов метилируется, и почти все - по 2'-гидроксильной группе рибозных остатков. Высокоспецифические участки метилирования сохраняются в рРНК таких эволюционно далеких эукариот, как дрожжи и плодовая мушка. Кроме того, более 100 остатков уридина изомеризуются в остатки псевдоуридина. Во время созревания происходит ассоциация множества рибосомных белков с этими РНК и их предшественниками. Возможно, именно взаимодействие РНК с рибосомными белками делает определенные участки чувствительными к действию нуклеаз.
Рис. 29.31. Образование рибосомной РНК млекопитающих из первичного транскрипта. Спей- серные участки показаны желтым
29.21. Информационные РНК избирательно образуются из больших ядерных предшественников РНК (гетерогенной ядерной РНК, гяРНК)
Образование рибосомных РНК у эукариот сходно с ее образованием у прокариот (разд. 25.17). В то же время между эукариотическими и прокариотическими мРНК имеются существенные различия.
1. Первичные транскрипты у эукариот не используются непосредственно в качестве мРНК. Прежде чем они переходят из ядра в цитозоль, они подвергаются процессингу. Трансляция и транскрипция у эукариот разобщены во времени и в пространстве, тогда как у прокариот они тесно сопряжены.
2. Первичные транскрипты у эукариот имеют в длину от 2 до 20 кb, в связи с чем их называют гетерогенной ядерной РНК (гяРНК). Обычно эти первичные транскрипты в несколько раз длиннее, чем мРНК, которые из них получаются. Образование эукариотических мРНК сопряжено со сплайсингом и расщеплением. До сих пор неизвестно, выполняет ли гяРНК еще какую-нибудь роль, кроме того, что она служит предшественником мРНК.
3. Эукариотические мРНК содержат на 5'-конце «колпачки» (кэпы), которые представляют собой модифицированные нуклеотиды. Кроме того, большинство мРНК несет на 3 -коние длинную poly(А)-последовательность.
4. Эукариотические мРНК моноци- стронны, т.е. они являются матрицами для синтеза только одной полипептидной цепи. Многие прокариотические мРНК, наоборот, полицистронны (например, мРНК лактозного оперона - матрица для синтеза трех полипептидных цепей).
5. Популяция молекул мРНК эукариотической клетки зависит не только от скорости транскрипции определенных генов; в эукариотических клетках имеется и еще один уровень принятия решений; должен ли тот или иной первичный транскрипт подвергнуться деградации или процессингу с образованием зрелой мРНК и последующим транспортом ее в цитозоль1.
1 Следует указать еще одно важное отличие прокариотических мРНК от эукариотических — время жизни. У прокариот оно составляет не более нескольких минут, а у эукариот - десятки минут или часы, а в исключительных случаях недели и месяцы. Прим. перев.
29.22. На 5'-конце мРНК находятся «колпачки», а на 3'-конце, как правило, poly(А)-последовательности
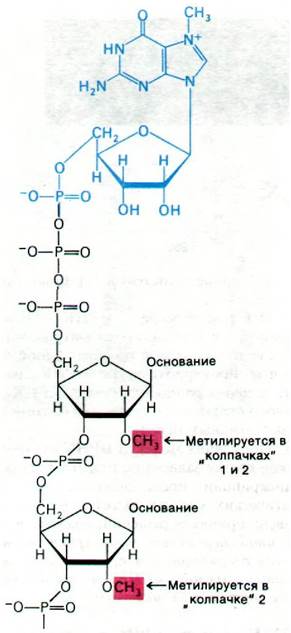
5'-конец всех известных эукариотических мРНК (но не тРНК или рРНК) модифицирован особым образом. К мРНК присоединен 7-метилгуанилат необычной пирофосфатной связью 5'—5' (рис. 29.32). Эта характерная структура называется «колпачком». Она присоединяется к первичному транскрипту. 5'-трифосфатный конец новообразованной цепи гидролизуется до дифосфата, и на него переносится остаток гуанилата GTP. Затем N-7 этого концевого гуанина метилируется S-аденозилметионином и образуется так называемый «колпачок» 0. Соседние остатки рибозы могут быть метилированы, при этом образуется «колпачок»1 или «колпачок» 2 (рис. 29.32). Эти «колпачки» способствуют стабилизации мРНК, защищая их 5'-концы от фосфатаз и нуклеаз. Кроме того, «колпачки» повышают эффективность трансляции мРНК в эукариотических системах синтеза белка.
1 В последнее время получен ряд данных, свидетельствующих об участии т.н. низкомолекулярных ядерных РНК в сплайсинге, а также в созревании З'-концов РНК. Эта фракция РНК исследуется уже давно. Она весьма консервативна и содержит последовательности, комплементарные участкам экзонов, прилежащим к цитронам. Низкомолекулярные ядерные РНК, по-видимому, обеспечивают узнавание участков сплайсинга соответствующими ферментами. - Прим. перев.
Рис. 29.32. Структура «колпачков», расположенных на 5'-конце эукариотических мРНК. Все «колпачки» содержат 7-метилгуанилат (изображен синим цветом), присоединенный пирофосфатной связью к 5'-концу. В «колпачке» 0 ни одна рибоза не метилирована, в «колпачке» 1 метилирована одна рибоза, в «колпачке» 2 - две
Кроме того, большинство эукариотических мРНК имеет на З'-конце poly(А)-последовательности. Роlу(А)-полимераза присоединяет 150-200 нуклеотидов к первичному транскрипту, на 3'-конце которого расположен динуклеотид GС, а на расстоянии примерно 20 нуклеотидов от него - последовательность AAUAA. Исследования различных мРНК, введенных в яйца Xenopus, показали, что роlу(А)-фрагмент увеличивает стабильность мРНК, но при этом не имеет никакого значения для трансляции. Кроме того, отсутствие роlу(А) на конце гистоновых мРНК показывает, что он не обязательно должен присутствовать в мРНК для ее транспорта из ядра в цитозоль.