БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 33. ИММУНОГЛОБУЛИНЫ
33.2. Синтез специфических антител в ответ на антиген
Животные способны синтезировать специфические антитела против практически любой химической группы. Особенно эффективно стимулирует образование антител динитрофенол (ДНФ), и потому его широко используют в качестве гаптенной детерминанты. Антитела против ДНФ получают следующим образом.
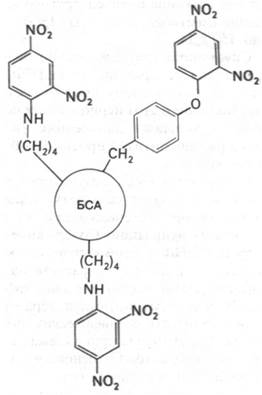
1. ДНФ-группы присоединяют ковалентно к белку-носителю, например, к альбумину сыворотки быка (БСА), используя взаимодействие фтординитробензола с боковыми цепями лизина или других нуклеофильных остатков белка (рис. 33.4).
Рис. 33.4. Динитрофенильное производное бычьего сывороточного альбумина (ДНФ-БСА) - эффективного антигена
2. ДНФ-БСА вводят кролику в качестве иммуногена (антигена). Спустя несколько дней содержание антител против ДНФ-альбумина начинает возрастать (рис. 33.5). Эти первыми появляющиеся антитела принадлежат к классу иммуноглобулинов М (IgM) массой около 1000 кДа.
Рис. 33.5. Кинетика появления иммуноглобулинов М и G (IgM и IgG) в сыворотке после иммунизации
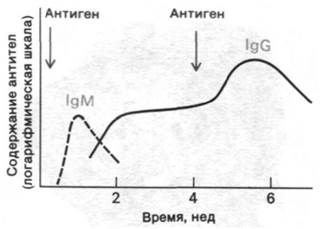
3. Спустя примерно 10 дней после введения антигена количество иммуноглобулина М начинает уменьшаться, но одновременно растет содержание антител другого класса, а именно иммуноглобулинов G (IgG) массой около 150 кДа.
4. Спустя примерно 3 недели после введения антигена содержание анти-ДНФ-антител класса иммуноглобулинов G достигает плато. Введение в этот период новой порции ДНФ-БСА вызывает дальнейшее увеличение содержания антител против ДНФ в сыворотке кролика.
5. Собирают кровь иммунизированного кролика и получают сыворотку (называемую антисывороткой, поскольку она получена после иммунизации). Содержание антител против ДНФ в этой сыворотке может достигать 1 мг антител в расчете на 1 мл сыворотки. ДНФ исключительно эффективен как агент, стимулирующий образование больших количеств специфических антител. Почти все эти антитела принадлежат к классу иммуноглобулинов G - основному классу иммуноглобулинов сыворотки.
6. Следующий этап - отделение антител против ДНФ от антител с другой специфичностью и от прочих сывороточных белков. Антитела против ДНФ отличаются от остальных белков антисыворотки очень высоким сродством к ДНФ. Следовательно, их можно выделить методом аффинной хроматографии. Для этого готовят колонку, содержащую динитрофенильные группы, ковалентно связанные с нерастворимым углеводным носителем. На колонку наносят антисыворотку и затем промывают буферным раствором. Большая часть белков антисыворотки проходит сквозь колонку, поскольку их сродство как к ДНФ-группам, так и к углеводному носителю ничтожно мало или вовсе отсутствует. В то же время антитела против ДНФ прочно связываются на колонке. Далее эти антитела снимают с колонки. Для этого на колонку наносят ДНФ в высокой концентрации: добавленный ДНФ связывается с антителами, вытесняя их из связи с динитрофенильными группами нерастворимого углеводного носителя. Растворимый комплекс, состоящий из ДНФ и антител против ДНФ, выходит с колонки. Далее ДНФ удаляют путем диализа или ионно-обменной хроматографии, и в результате остается препарат очищенных антител.
33.3. Участки антител, связывающие антиген, подобны активным центрам ферментов
Участки связывания антигена в молекулах антител во многих отношениях сходны с активными центрами ферментов.
1. Константы связывания для гаптенов были определены методами равновесного диализа и спектроскопии. Например, присоединение окрашенного гаптена типа динитрофенильного производного тушит флуоресценцию остатков триптофана в белке-антителе. Степень тушения служит мерой насыщения участков связывания в молекуле белка. Для большинства гаптенов константы связывания лежат в пределах от 10-4 до 10 10 М. Следовательно, стандартная свободная энергия связывания составляет от —6 до — 15 ккал/моль, т. е. лежит в границах, характерных для фермент-субстратных и фермент-коферментных комплексов. Кроме того, комплексы гаптен-антитело образуются под действием тех же сил, что и фермент-субстратные комплексы. Сочетание слабых нековалентных связей типа электростатических, водородных и вандерваальсовых обеспечивает прочное и специфическое связывание.
2. Проверяли способность антител к декстрану (полисахариду, состоящему из остатков глюкозы) связывать олигомеры. Обнаружили, что полное связывающее сродство проявляется в отношении олигомера, состоящего из шести остатков глюкозы. Здесь напрашивается сравнение с лизоцимом, у которого в щели активного центра размещается тоже шесть углеводных остатков. Из этого сопоставления следует, что длина связывающего участка в молекулах антител против декстрана составляет 25 А.
3. На основе спектроскопических свойств ряда гаптенов можно получить информацию о степени полярности участков связывания в молекулах антител. Так, некоторые нафталины, находясь в высокополярном окружении (например, в воде), дают слабую желтую флуоресценцию, а в выраженно неполярном окружении (например, в гексане)- интенсивно голубую. Если такой нафталиновый гаптен присоединяется к специфическому антителу, то появляется интенсивно голубая флуоресценция, что указывает на изгнание воды из участка связывания. Как правило, участки связывания представляют собой неполярные ниши в молекуле антитела; это увеличивает прочность связывания антигена по причинам, которые уже обсуждались применительно к фермент- субстратному взаимодействию (разд. 6.8).
4. Гаптен довольно точно соответствует связывающему участку по структуре. Специфичность связывания, безусловно, очень высока, но не абсолютна. Гаптен прочно удерживается в участке связывания, имея, таким образом, малую свободу вращения. Константа скорости связывания для многих гаптенов составляет около 108 М-1 • с-1.
Столь высокая константа свидетельствует о том, что скорость процесса контролируется скоростью диффузии. Связывание гаптена не сопровождается, по-видимому, существенными структурными перестройками.
33.4. Препараты антител с определенной специфичностью обычно гетерогенны
Антитела имеют существенное отличие от ферментов: большинство нормальных антител с определенной специфичностью, например, антитела против ДНФ, неоднородны по молекулярному составу. Анализ связывания динитрофенильных гаптенов с препаратом анти-ДНФ-антител выявил целый набор величин сродства. Некоторые молекулы антител связывают ДНФ с К = 10-6 М, тогда как другие - с К = 10-10 М. В отличие от этого ферменты, как правило, характеризуются только одной константой связывания данного субстрата или кофермента. Далее при электрофорезе препарата антител против ДНФ или других специфических антител выявляется множество полос белка. Ферменты же при электрофорезе дают одну полосу или небольшое число отдельных полос (например, изоферменты лактат-дегидрогеназы).
Гетерогенность антител с данной специфичностью определяется самой природой иммунного ответа. В чем причина гетерогенности? Оказалось, что антитела, продуцируемые одной клеткой, гомогенны. Однако различные клетки образуют разные по структуре антитела. Антитела против ДНФ синтезируются большим количеством различных клеток, что и обусловливает их гетерогенность.