БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 24 ДНК: ГЕНЕТИЧЕСКАЯ РОЛЬ, СТРУКТУРА И РЕПЛИКАЦИЯ
24.19. Одна цепь ДНК синтезируется прерывисто
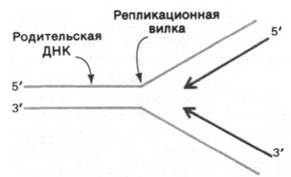
Вернемся к взаимодействию молекул при репликации. В области репликационной вилки обе цепи родительской ДНК служат матрицами для синтеза новой ДНК. Напомним, что цепи родительской ДНК антипараллельны. Следовательно, общее направление синтеза ДНК должно быть 5' → 3' для одной из дочерних цепей и 3' → 5' для другой (рис. 24.39). Однако все известные ДНК-полимеразы синтезируют ДНК в направлении 5' → 3', а не 3' → 5'. Как же тогда происходит кажущийся (при низкой разрешающей способности метода) рост одной из дочерних цепей в направлении 3' → 5'?
Рис. 24.39. При низком разрешении кажущееся направление репликации ДНК будет 5' → 3' для одной дочерней цепи и 3' → 5' для другой. На самом деле обе цепи синтезируются в направлении 5' → 3', как показано на рис. 24.40
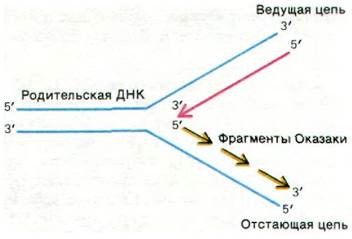
Проблема была решена Рейдзи Оказаки (Reiji Okazaki), который обнаружил, что значительная часть новосинтезированной ДНК существует в виде коротких фрагментов. Такие фрагменты длиной около 1000 нуклеотидов (они называются фрагментами Оказаки) существуют в течение непродолжительного времени в непосредственной близости от репликационной вилки. По мере прохождения репликации эти фрагменты соединяются друг с другом ковалентно под действием ДНК-лигазы и образуют одну из дочерних цепей (рис. 24.40). Другая новая цепь синтезируется непрерывно или почти непрерывно. Та цепь, которая образуется из фрагментов Оказаки, называется отстающей цепью, а та, что синтезируется без разрывов или почти без разрывов, ведущей цепью. И фрагменты Оказаки, и ведущая цепь синтезируются в направлении 5' → 3'. Прерывистая сборка отстающей цепи позволяет путем полимеризации в направлении 5' → 3' на атомном уровне получать общий рост цепи в направлении 3' → 5'.
Рис. 24.40. Схематическое изображение репликационной вилки. Обе цепи ДНК синтезируются в направлении 5' → 3'. Ведущая цепь синтезируется непрерывно, а отстающая - в виде коротких фрагментов (фрагменты Оказаки)
24.20. Затравкой синтеза ДНК служит РНК
Как начинается синтез ДНК? Напомним, что всем ДНК-полимеразам для инициирования синтеза ДНК необходима затравка со свободной З'-ОН-группой. Что служит затравкой при синтезе ведущей цепи и фрагментов Оказаки? Важным толчком к решению этого вопроса послужило наблюдение, что для инициации синтеза ДНК необходим синтез РНК. На основе этого открытия было высказано предположение, что РНК, очевидно, служит затравкой в синтезе ДНК, так как уже было известно, что РНК-полимеразы способны начинать синтез цепей de novo. Затем было показано, что новообразующаяся ДНК ковалентно связана с коротким фрагментом РНК, который и служит затравкой. Итак, РНК-затравка в синтезе ДНК.
По всей вероятности, репликация ДНК в клетке E. coli происходит, как показано на рис. 24.41.
Рис. 24.41. Инициация синтеза ДНК. А - праймаза синтезирует короткую комплементарную цепь РНК; Б - эта РНК служит затравкой для синтеза новой ДНК; В - РНК, входящая в состав новообразованной цепи, гидролизуется, при этом образуется брешь, которая впоследствии заполняется
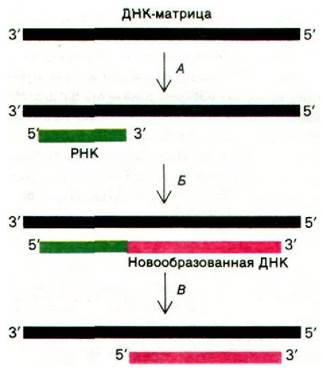
1. Особая РНК-полимераза (ее называют праймаза) синтезирует короткую цепь РНК (примерно 10 нуклеотидов), комплементарную одной из цепей ДНК-матрицы. В отличие от ДНК-полимеразы праймаза не нуждается в затравке для синтеза полинуклеотида.
2. 3'-гидроксильная группа концевого рибонуклеотида этой цепи РНК служит затравкой для синтеза ДНК под действием голофермента ДНК-полимеразы III. Большая часть новообразованной ДНК синтезируется этим мультисубъединичным комплексом.
3. РНК-компонент этого РНК-ДНК-гибрида гидролизуется под действием ДНК- полимеразы I.
4. После удаления РНК из новообразованных цепей между фрагментами ДНК остаются довольно обширные бреши. ДНК- полимераза I, которая хорошо приспособле-
на для синтеза ДНК на одноцепочечной матрице, заполняет эти бреши.
Последние исследования показали, что действию праймазы предшествует образование предзатравочного промежуточного комплекса, состоящего как минимум из пяти белков. Один из них - белок dnaB - может передвигаться вдоль ДНК, используя энергию гидролиза АТР. Белок dnaB может служить сигналом для активации праймазы. Специфичность инициации очередного цикла репликации может обеспечиваться белками, доставляющими белок dnaB точно в область гена ilv, где находится точка начала репликации хромосомы E. coli. Время начала репликации ДНК имеет критическое значение, поскольку оно должно быть скоординировано с делением клетки. И действительно, бактериальная хромосома ассоциирована с впячиванием клеточной мембраны (рис. 24.42).
Рис. 24.42. Электронная микрофотография хромосомы Е. coli, прикрепленной к двум фрагментам клеточной мембраны. На рисунке изображена одна интактная суперспирализованная молекула ДНК

24.21. Энергия гидролиза АТР используется для расплетания родительской ДНК в области репликационной вилки под действием белка rep
В 1953 г. Уотсон и Крик отмечали, что «раскрутить спираль - задача труднопреодолимая». Исследования, проведенные в последнее время, показали, что в клетке E. coli в области репликационной вилки происходит активное расплетание родительской двойной спирали под действием фермента — белка rep. Поскольку энергия для расплетания родительской ДНК высвобождается при гидролизе АТР, белок гер называют хеликазой. На разделение каждой пары оснований затрачиваются примерно две молекулы АТР. Затем каждая из разделенных цепей родительской ДНК взаимодействует с несколькими молекулами белка, связывающегося с одноцепочечной ДНК (ОЦ-связывающий белок). Роль ОЦ-связывающего белка - стабилизировать одноцепочечные участки ДНК, образовавшиеся под действием хеликазы, чтобы расплетенная область могла функционировать в качестве матрицы. ОЦ-связывающие белки называют также белками, дестабилизирующими спираль (белки ДС), или плавящими белками.
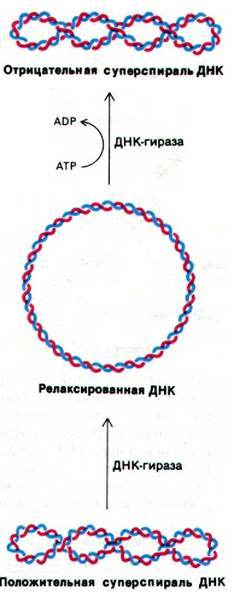
24.22. ДНК-гираза вводит отрицательные супервитки в родительскую ДНК, чтобы облегчить ее расплетание
При расплетании ковалентно замкнутой кольцевой молекулы ДНК возникают топологические проблемы, так как расплетание двойной спирали вызывает образование положительных супервитков в замкнутой молекуле. В области репликационной вилки родительская ДНК вращается со скоростью 100 об/с - более чем в 100 раз быстрее, чем обычная долгоиграющая пластинка. Чтобы процесс расплетания продолжался, необходимо снять эти положительные супервитки, образовавшиеся в результате вращения. Другими словами, необходимо наличие какого-то молекулярного шарнира. Недавно Мартин Геллерт (Martin Gellert) установил, что эту функцию выполняет ДНК-гираза. Эта топоизомераза удаляет положительные супервитки, внося одноцепочечные разрывы и затем заделывая фосфодиэфирные связи в остове ДНК. АТР не требуется для такой термодинамически выгодной релаксации третичной структуры ДНК. Более того, ДНК-гираза может активно вводить отрицательные супервитки в ковалентно замкнутую кольцевую ДНК за счет энергии гидролиза АТР (рис. 24.43). Эти отрицательные супервитки способствуют расхождению цепей родительской ДНК в области репликационной вилки1.
1 В этой главе автор называет ДНК-гиразой два совершенно различных фермента. Один из них - ДНК-гираза, способная вводить в ДНК термодинамически невыгодные отрицательные супервитки за счет энергии гидролиза АТР; другой - независимая от АТР ДНК-топоизомераза, приводящая кольцевую молекулу ДНК в термодинамически равновесное (релаксированное) состояние. Это два совершенно различных белка: у них различные ингибиторы, потребности в ионных условиях среды, механизм действия. В репликации в качестве молекулярного шарнира участвует ДНК-гираза. Прим. перев.
Рис. 24.43. Каталитические активности ДНК-гиразы