БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 34. МЫШЕЧНОЕ СОКРАЩЕНИЕ И ПОДВИЖНОСТЬ КЛЕТОК
34.16. Цитохалазин и фаллоидин тормозят подвижность, сопряженную с процессами сборки и дезагрегации нитей актина
Цитохалазин В (алкалоид из грибов) при добавлении к эукариотическим клеткам изменяет их форму и подавляет многие формы подвижности. Так, этот алкалоид ингибирует образование складчатых краев у фибробластов, отрастание аксонов от ганглиев, ретракцию кровяного сгустка тромбоцитами, деление оплодотворенной яйцеклетки морского ежа. Судя по данным электронной микроскопии, цитохалазин оказывает влияние на микрофиламенты, так как они исчезают в клетках, обработанных этим алкалоидом. Оказалось, что цитохалазин препятствует сборке актиновых нитей, специфически взаимодействуя с одним из концов нити. Этот ингибиторный эффект цитохалазина свидетельствует о динамичности структуры микрофиламентов.
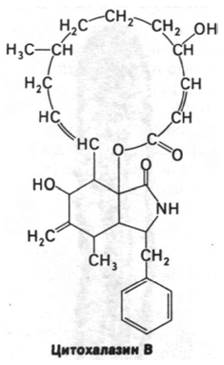
Значение постоянно идущих процессов сборки и дезагрегации микрофиламентов для клеточного движения было выявлено также при изучении механизма действия фаллоидина. Этот токсический агент присутствует в ядовитом грибе Amanita phalloides, содержащем также а-аманитин — ингибитор РНК-полимеразы (разд. 29.18). Подобно цитохалазину, фаллоидин блокирует те формы подвижности, в которых участвуют микро- филаменты. Фаллоидин присоединяется к единицам актина в микрофиламентах и тем самым препятствует его деполяризации. Таким образом, фаллоидин как бы запирает микрофиламенты.
34.17. Микротрубочки участвуют в различных видах клеточной подвижности и частично формируют цитоскелет
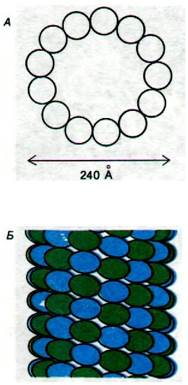
До сих пор мы рассматривали роль микрофиламентов в различных видах клеточной подвижности. Обратимся теперь к другому классу волокнистых элементов, а именно к микротрубочкам, которые имеются практически во всех эукариотических клетках и участвуют как в поддержании архитектуры клетки, так и в сократительной активности (рис. 34.29). Микротрубочки представляют собой полые цилиндрические структуры, образованные из двух сходных (массой по 55 кДа) типов субъединиц - α и β-тубулина. Наружный диаметр микротрубочек составляет около 240 А, и этим они четко отличаются от микрофиламентов (диаметром 70 А) и от промежуточных филаментов (структур диаметром 100 А, выполняющих роль связывающих элементов). Жесткая стенка микротрубочек образована из спирально уложенных, чередующихся α- и β-тубулиновых субъединиц (рис. 34.30). Повторяющейся единицей структуры является αβ-димер, и в целом структура образована из 13 протофиламентов, идущих параллельно длинной оси микротрубочки.
Рис. 34.29. Иммунофлуоресцентная микрофотография, демонстрирующая распределение микротрубочек в фибробласте
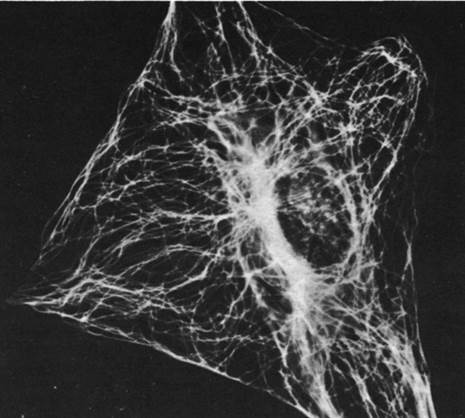
Рис. 34.30. Спиральное расположение тубулиновых субъединиц в микротрубочке. А - схематическое изображение поперечного среза; видно, что микротрубочка образована 13 протофиламентами. Б - поверхностная решетка из α- и β-субъединиц
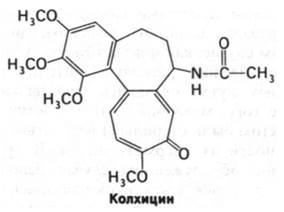
Сборка и дезагрегация микротрубочек происходят с противоположных концов. Алкалоид из осенних крокусов колхицин блокирует сборку и тем самым тормозит клеточные процессы, протекающие с участием микротрубочек. Например, колхицин останавливает клеточное деление на стадии метафазы, так как микротрубочки имеют существенное значение для расхождения хромосом. Колхицин ингибирует также направленное движение разного рода частиц в клетках, что лежит в основе торможения процессов их секреции. На протяжении нескольких веков колхицин используют для лечения острых приступов подагры.
34.18. Биение ресничек и движение жгутиков обусловлено скольжением микротрубочек, индуцированным динеином
Микротрубочки являются основными компонентами ресничек и жгутиков у эукариот. Эти органеллы имеются во многих клетках. Подвижные реснички, подобно веслам, вызывают ток жидкости параллельно поверхности клетки. Так, координированное движение ресничек, выстилающих дыхательные пути, способствует удалению чужеродных частиц. Жгутики необходимы для продвижения свободных клеток, таких, как спермии или простейшие. Как показали электронномикроскопические исследования, структура ресничек и жгутиков практически у всех эукариот в основе своей одинакова. Это пучок волокон, называемый аксонемой, который окружен мембраной, составляющей продолжение плазматической мембраны. Волокна в аксонеме - это микротрубочки. По периферии расположены девять двойных микротрубочек, в центре - две одинарные микротрубочки (рис. 34.31). Такой часто встречающийся структурный мотив известен как расположение 9 + 2. Диаметр двух центральных микротрубочек равен 240 А. Каждая из девяти периферических пар на поперечном срезе имеет вид цифры 8 и размер 370 х 250 А (рис. 34.32). Одна из микротрубочек в паре - субфибрилла А - меньше по размеру и присоединяется к центральной капсуле реснички с помощью радиального мостика. От каждой субфибриллы А отходят две ручки. В отдельной ресничке все ручки направлены в одну сторону. Обработка ресничек детергентом и далее раствором солей высокой концентрации приводит к удалению наружной мембраны и солюбилизации АТРазы, называемой динеином. В результате этой обработки расположенные на периферии волокна сохраняют свою цилиндрическую попарную организацию, но утрачивают ручки. При добавлении динеина в соответствующей ионной среде ручки восстанавливаются. Следовательно, состоящие из динеина ручки субфибриллы А обладают АТРазной активностью.
Рис. 34.31. Электронная микрофотография поперечного среза аксо- немы жгутика. Девять наружных двойных микротрубочек окружают две одинарные
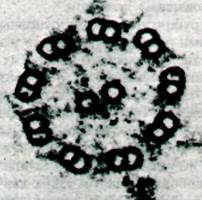
Рис. 34.32. Схематическое изображение структуры аксонемы
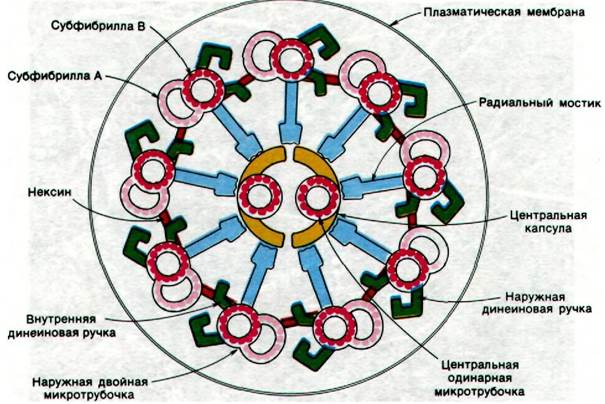
Каким образом расщепление АТР динеи- ном вызывает движение ресничек и жгутиков? Как показали Питер Сатир и Ян Гиббонс (Peter Satir, Jan Gibbons), периферические двойные микротрубочки аксонемы скользят относительно друг друга и тем самым вызывают изгиб реснички. Само скольжение обусловлено образованием ди- неиновых поперечных мостиков. По-видимому, динеиновые ручки одной пары микротрубочек по мере гидролиза АТР продвигаются вдоль прилегающей пары совершенно аналогично тому, как в скелетной мышце миозиновые поперечные мостики движутся вдоль актиновой нити. В интактной ресничке радиальные мостики противодействуют скольжению, и благодаря им происходит не сокращение реснички, а локальный изгиб.
При обследовании группы больных хроническими легочными заболеваниями были обнаружены аксонемы, лишенные динеина. В этом случае, как показал Бьёрн Афзелиус (BjornAfzelius), реснички эпителия дыхательных путей оказались неподвижными. Более того, мужчины с этим генетическим дефектом были стерильны вследствие неподвижности их сперматозоидов. В другом, недавно обнаруженном случае неподвижность ресничек и жгутиков оказалась обусловленной дефектом радиальных мостиков. Эти клинические наблюдения подтверждают существующие представления о механизме движения рассматриваемых органелл.