БІОХІМІЯ ДЛЯ ВЧИТЕЛЯ - Ф.Ф. БОЄЧКО - 1985
ЕНЕРГЕТИКА БІОЛОГІЧНИХ ПРОЦЕСІВ
ПЕРЕТВОРЕННЯ ВУГЛЕВОДІВ У КЛІТИНАХ
Як відомо, вуглеводи становлять основну масу рослин (близько 75—85 % У перерахунку на суху масу) і велику частину харчового раціону людини, а також травоїдних тварин. Вуглеводи виконують в організмі важливу пластичну функцію. Вони є джерелом органічних сполук, які використовуються в біосинтезі білків, ліпідів та ряду інших важливих речовин. У процесі розщеплення вуглеводів вивільнюється значна кількість енергії, яка необхідна для нормальної життєдіяльності організму. Організм людини за рахунок вуглеводів одержує 60—70 % енергії.
Перетворення вуглеводів у тканинах людини і тварин відбуваються анаеробно (при відсутності кисню) і аеробно (при наявності кисню). У першому випадку кінцевим продуктом є молочна кислота:
![]()
При аеробному перетворенні вуглеводів кінцевими продуктами є оксид вуглецю (IV) і вода:
![]()
Анаеробне перетворення вуглеводів
Гліколіз. Цей вид перетворення вуглеводів виник раніше, ніж аеробне перетворення вуглеводів. Лише після того, як в атмосфері Землі з’явився кисень, виникло аеробне перетворення. Утворення вільного кисню тісно пов’язане з процесом фотосинтезу.
Основним вуглеводом, що міститься в клітинах і використовується в процесі анаеробного перетворення, є глюкоза. Основні етапи цього процесу уже з’ясовано. Глюкоза, перед тим як включитись у процес перетворення, піддається фосфорилюванню з участю АТФ, в результаті чого утворюється глюкозо-6-фосфат. Якщо джерелом глюкози є полісахариди, то спочатку відбувається їх фосфорилювання з участю неорганічного фосфату. Глюкозо-1-фосфат, що утворився внаслідок ізомеризації, перетворюється в глюкозо-6-фосфат. Ця сполука утворюється і при використанні як джерела глюкози інших цукрів, зокрема дисахаридів.
Глюкозо-6-фосфат внаслідок ряду перетворень розпадається на дві молекули фосфотріоз — гліцеринальдегід-3-фосфат і діоксіацетонфосфат. Останній перетворюється в гліцеринальдегід-3-фосфат. Отже, з однієї молекули глюкози утворюються дві молекули гліцеринальдегід-3-фосфату.
На наступних етапах гліцеринальдегід-3-фосфат перетворюється в молочну кислоту (лактат). Ці етапи перетворень є спільними для всіх цукрів. Вони охоплюють окислювально-відновні реакції і механізми нагромадження енергії за рахунок фосфорилювання АДФ та утворення АТФ.
Реакції гліколізу каталізують одинадцять різних ферментів. Кожний з них уже добре вивчено і добуто в чистому вигляді. При цьому більшу частину цих ферментів добуто також і в кристалічному вигляді. Усі ферменти гліколізу добре розчинні у воді, і кожний з них каталізує свою реакцію, незалежно від інших ферментів цього процесу. Це свідчить про те, що ферменти гліколізу не зв’язані з клітинними компонентами і перебувають у цитоплазмі в розчиненому стані. Вони не утворюють таких поліферментних систем, як дихальний ланцюг реакцій окислювального фосфорилювання.
У процесі реакцій гліколізу з однієї молекули гексози утворюються дві молекули фосфотріоз. Кожна з них у процесі перетворення до молочної кислоти зумовлює утворення двох молекул АТФ, тобто всього утворюється чотири молекули АТФ. Оскільки дві молекули АТФ використовуються для фосфорилювання глюкози і фруктозо-6-фосфату, то чистий вихід енергії дорівнюватиме двом молекулам АТФ, у макроергічних зв’язках яких акумулюється близько 84 кДж енергії. Отже, загальне сумарне рівняння гліколізу можна показати так:
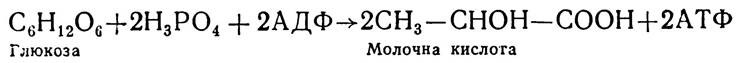
Енергетично процес гліколізу для організму невигідний, бо, щоб перекрити енергетичні затрати, потрібна велика кількість вуглеводів. Та як фізіологічний процес він дуже важливий для організму, тому що організм може виконувати різноманітні види робіт в умовах недостатнього постачання його киснем.
Спиртове бродіння. Процес спиртового бродіння, який здійснюється мікроорганізмами (переважно дріжджами), за хімізмом дуже близький до гліколізу. Усі стадії перетворення вуглеводів до піровиноградної кислоти в обох випадках відбуваються однаково. Відмінність між ними починається після етапу утворення піровиноградної кислоти:
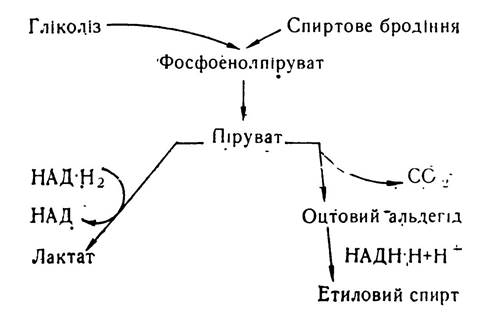
При гліколізі піровиноградна кислота з участю ферменту НАД-залежної дегідрогенази відновлюється і перетворюється в молочну кислоту. В умовах спиртового бродіння піровиноградна кислота з участю ферменту піруватдекарбоксилази декарбоксилюється з утворенням оцтового альдегіду:
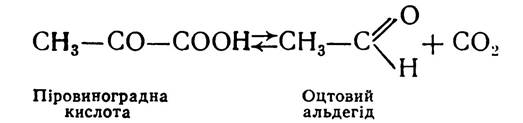
Потім з участю НАД·H+Н+ - залежної алкогольдегідро генази оцтовий альдегід відновлюється до етилового спирту:
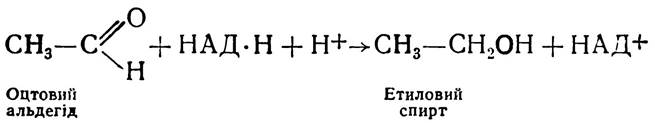
Отже, етиловий спирт і оксид вуглецю (IV) є кінцевими продуктами спиртового бродіння. Сумарне рівняння спиртового бродіння можна показати так:
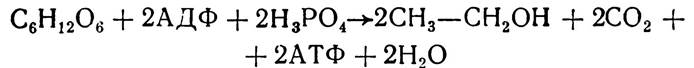
Реакції, в результаті яких відбувається нагромадження енергії у вигляді АТФ, для процесів спиртового бродіння такі самі, як і для гліколізу.
Крім спиртового бродіння, відомі й інші види бродіння, кінцевими продуктами яких є пропіонова, масляна і янтарна кислоти та ацетон.
Аеробне перетворення вуглеводів
Основним джерелом енергії для організму людини, тварин і рослин є аеробне перетворення вуглеводів. Між анаеробним та аеробним розщепленням вуглеводів існує тісний взаємозв’язок. Насамперед він виявляється в наявності спільної зв’язуючої ланки — піровиноградної кислоти. Ця кислота в анаеробних умовах піддається відновленню і перетворюється до молочної кислоти, яка є кінцевим продуктом гліколізу. В аеробних умовах піровиноградна кислота з участю цілого ряду коферментів — тіамінпірофосфату (ТПФ), ліпоєвої кислоти (ЛК), коензиму А (КоА), НАД+ і ФАД — перетворюється до ацетил-КоА:
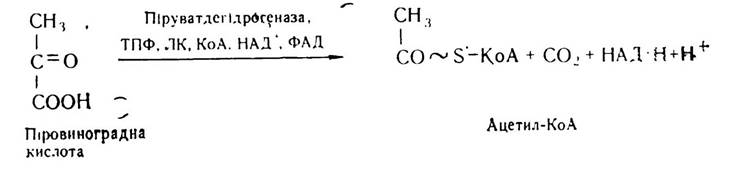
Частина молекули цієї сполуки використовується для синтезу жирів, вуглеводів та інших сполук, частина вступає в цикл три- і дикарбонових кислот і окислюється до кінцевих продуктів — Н2O і СО2.
Цикл три- і дикарбонових кислот, його ще називають циклом Кребса, оскільки окислення органічних сполук за допомогою такого механізму вперше відкрив англійський дослідник Кребс.
Цей цикл складається з восьми послідовних реакцій (див. форзац II). Починається цикл Кребса з конденсації ацетил-КоА з щавлевооцтовою кислотою, в результаті чого утворюється лимонна кислота. Потім лимонна кислота (шестивуглецева сполука) завдяки дегідруванню і декарбоксилюванню втрачає (відщеплює) два атоми вуглецю і знову в циклі Кребса з’являється чотиривуглецева сполука — щавлевооцтова кислота. Отже, внаслідок повного обороту циклу молекула ацетил-КоА згоряє (окислюється) до СO2 і Н2O:
![]()
Для безперервного функціонування циклу необхідне систематичне надходження в його систему ацетил-КоА, а коферменти НАД+ і ФАД, які перейшли у відновлену форму, повинні постійно окислюватись. Це окислення відбувається в ланцюгу дихальних ферментів, який зосереджений в мітохондріях.
Енергія, що вивільнюється внаслідок окислення оцтової кислоти, нагромаджується в макроергічних зв’язках АТФ.
Як бачимо, у циклі Кребса утворюються дві молекули НАД·Н+Н+, одна молекула НАДФ·Н+Н+ і одна молекула ФАД·Н2. Отже, всього утворюється чотири молекули відновлених коферментів. Доведено, що внаслідок окислення однієї молекули НАД·Н+Н+ або НАДФ·Н+Н+ відщепленням водню в ланцюгу дихальних ферментів синтезуються три молекули АТФ. Внаслідок окислення ФАД-Н2 утворюються дві молекули АТФ і одна молекула ГТФ (АТФ) на рівні субстрату при перетворенні сукциніл-КоА в янтарну кислоту. Отже, при перетворенні однієї молекули ацетил-КоА в циклі три- і дикарбонових кислот синтезується 3-3+3=12 молекул АТФ.
Слід зазначити, що три молекули АТФ синтезуються внаслідок окислення однієї молекули НАД·Н+Н+, яка утворюється при перетворенні піровиноградної кислоти до ацетил-КоА. Таким чином, при перетворенні однієї молекули піровиноградної кислоти до ацетил-КоА, а останнього — до СO2 і Н2O, синтезується 12+3=15 молекул АТФ. Оскільки з однієї молекули глюкози утворюються дві молекули піровиноградної кислоти, то всього утворюється 15·2 = 30 молекул АТФ.
Крім того, шість молекул АТФ синтезуються внаслідок окислення двох молекул НАД·Н2, що утворюються при перетворенні 3-фосфогліцеринового альдегіду, і дві молекули АТФ утворюються в процесі гліколізу. Отже, загальний енергетичний баланс перетворення однієї молекули глюкози до СO2 і Н2O становитиме 30+8=38 молекул АТФ, у макроергічних зв’язках яких нагромаджується 38·42=1596 кДж енергії. Перетворення однієї молекули глюкози в анаеробних умовах дає лише дві молекули АТФ, в макроергічних зв’язках яких вуге 2·42 = 84 кДж енергії. Отже, основним джерелом енергії для організму є аеробне окислення біологічних субстратів.
Біосинтез вуглеводів
Процес біосинтезу глюкози та інших вуглеводів є досить важливим у живій природі. Фотосинтезуючі організми, або автотрофи, утворюють величезну кількість гексоз з вуглекислого газу і води, застосовуючи у вигляді енергії сонячне світло. Такі гексози використовуються для утворення крохмалю, клітковини та інших полісахаридів,
У процесах асиміляції перетворення піровиноградної кислоти в глюкозу та інші вуглеводні компоненти клітини є центральним метаболічним шляхом, спільним для більшості представників тваринного світу і людини. У хребетних найбільш інтенсивно глюконеогенез відбувається в клітинах печінки і нирок (корковий шар).
Необхідно зазначити, що більшість стадій глюконеогенезу — це зворотні реакції гліколізу. Тільки три реакції гліколізу — гексокіназна, фосфофруктокіназна і фосфопі- руваткіназна — є незворотними. Тому на цих трьох етапах використовуються інші, обхідні шляхи, які енергетично більш сприятливі для проходження біосинтезу.