БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 10. МЕТАБОЛІЗМ НУКЛЕЇНОВИХ КИСЛОТ
10.5. Біосинтез нуклеотидів
10.5.2. Біосинтез піримідинових нуклеотидів
Біосинтез піримідинових нуклеотидів de novo. Піримідинові нуклеотиди, як і пуринові, в основному синтезуються з простих попередників de novo - СО2, глутаміну та аспарагінової кислоти (рис. 10.11), і тільки 10-20 % від загальної кількості - шляхом реутилізації з азотистих основ або нуклеозидів.
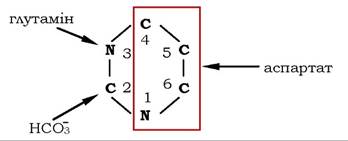
Рис. 10.11. Джерела атомів піримідинів
Біосинтез піримідинових нуклеотидів de novo відрізняється від синтезу пуринових тим, що піримідинове кільце спочатку формується, а вже потім до нього приєднується рибозофосфат.
Синтез піримідинового кільця (рис. 10.12) відбувається в цитозолі за участю карбамоїлфосфату, який утворюється з глутаміну, СО2 та АТФ у реакції, що каталізується карбамоїлфосфатсин- тазою ІІ (КФС ІІ). Доречно зазначити, що в процесі синтезу сечовини в мітохондріях за участю карбамоїлфосфатсинтетази І (КФС І) теж утворюється карбамоїлфосфат, але із СО2, АТФ та NH3. На метаболічному шляху синтезу піримідинових нуклеотидів активність цитозольного регуляторного ферменту КФС ІІ гальмується кінцевим продуктом - УМФ; на активність мітохондріального ферменту КФСІ УМФ не впливає. Таким чином, особливості внутрішньоклітинного розподілу ферментів і регуляторних механізмів дозволяють процесам синтезу піримідинових нуклеотидів і сечовини функціонувати незалежно й паралельно один одному, відповідно до потреб організму.
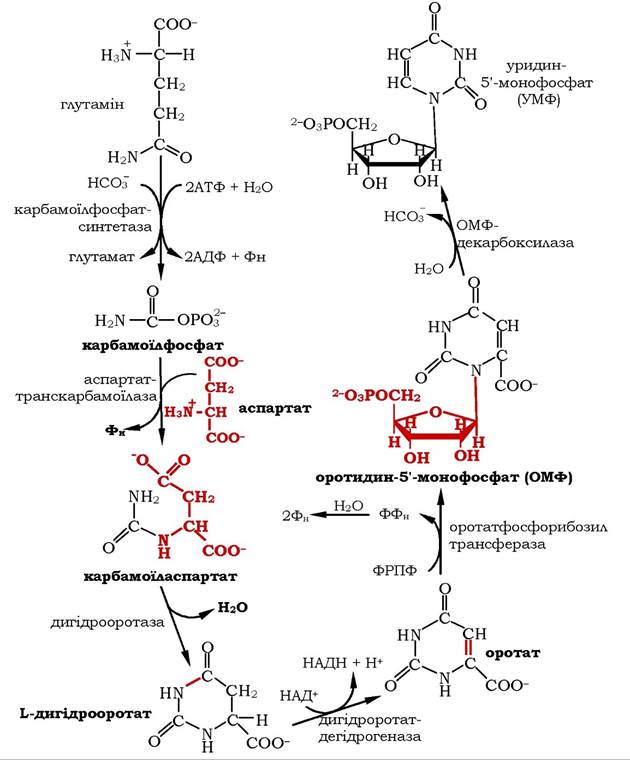
Рис. 10.12. Синтез уридин-5'-монофосфату
На наступному етапі цитоплазматичний карбамоїлфосфат конденсується з аспарагіновою кислотою під дією аспартаткарбамоїлази, а потім у реакції, що каталізується дигідрооротазою, відокремлюється Н2О й утворюється піримідинове кільце - дигі- дрооротова кислота. Дегідрування останньої до оротової кислоти відбувається за наявності ферменту дигідрооротатдегідрогенази з використанням НАД як коферменту.
Далі до оротової кислоти приєднується залишок рибозо-5- фосфату від 5-фосфорибозил-1-пірофосфату(ФРПФ). У результаті утворюється нуклеотид - оротидин-5'-монофосфат(ОМФ). Перетворення оротату в ОМФ каталізується оротатфосфорибозилтрансферазою.
Перший справжній піримідиновий рибонуклеотид - уридин-5'- монофосфат (УМФ) утворюється за дії ферменту ОМФ-декарбоксилази шляхом декарбоксилювання ОМФ. УМФ є попередником усіх інших піримідинових нуклеотидів.
Біосинтез УДФ, УТФ і цитидилових нуклеотидів. Фосфо- рилювання піримідинових нуклеозидмонофосфатів до відповідних ди- та трифосфатів відбувається за участю специфічних нуклеозидмонофосфат- і нуклеозиддифосфаткіназ шляхом перенесення γ-фосфатного залишку АТФ на відповідний субстрат:
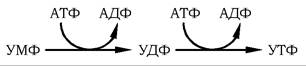
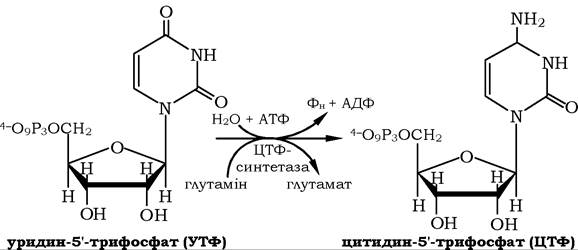
Перетворенню уридилової кислоти на цитидиловий нуклеотид має передувати фосфорилювання УМФ до уридин-5'-трифосфа- ту. За участю ЦТФ-синтетази відбувається амідування УТФ шляхом АТФ-залежного заміщення кетогрупи на амідну групу глута- міну з утворенням цитидин-5'-трифосфату:
Утворення третього піримідинового нуклеотиду - дТТФ - буде наведено нижче.
Синтез піримідинових нуклеотидів шляхом реутилізації.
Крім указаного ланцюга реакцій синтезу піримідинових нуклеотидів de novo, може мати місце пряме включення вільних піримідинових основ і нуклеозидів до складу нуклеотидів. Так, уридинфосфорилаза каталізує реакцію рибозилювання урацилу з утворенням уридину:
урацил + рибозо-1-фосфат →уридин + Н3РО4.
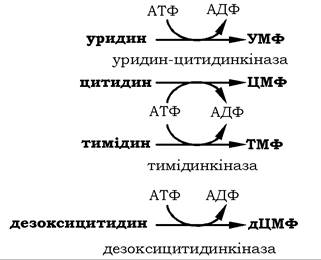
Утилізація піримідинових рибонуклеозидів у клітинах ссавців відбувається ефективніше, ніж окремих азотистих основ. Нижче наведено можливі реакції утворення піримідинових нуклеозидмонофосфатів з відповідних піримідинових нуклеозидів, які каталізуються нуклеозидкіназою:
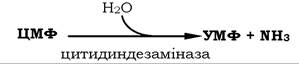
Завдяки гідролітичному дезамінуванню частина утвореного шляхом реутилізації ЦМФ може перетворюватися в УМФ під дією цитидиндезамінази й поповнювати запаси уридилових нуклеотидів:
Регуляція синтезу піримідинових нуклеотидів. Шлях синтезу піримідинових нуклеотидів контролюється за допомогою механізмів, які регулюють активність ферментів за типом зворотного зв'язку. Так, аспартаттранскарбамоїлаза досить чутлива до інгібуючої дії ЦТФ, але активується АТФ. Карбамоїлфосфатсинтетазна активність алостерично інгібується УМФ і УТФ, пуриновими нуклеотидами (рис. 10.13).
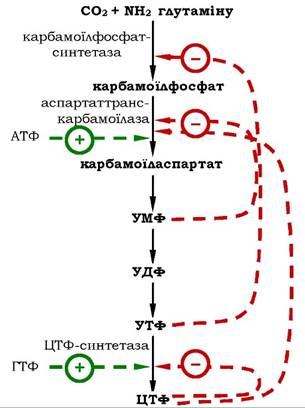
Рис. 10.13. Схема регуляції синтезу піримідинових нуклеотидів
Активність ЦТФ-синтетази, яка каталізує реакцію утворення ЦТФ із УТФ, гальмується власним продуктом - ЦТФ, але активується ГТФ.
Подібний спосіб регуляції дозволяє запобігти надлишковому синтезу не тільки УМФ, але й інших піримідинових нуклеотидів і забезпечити збалансоване утворення всіх чотирьох основних пуринових і піримідинових нуклеотидів, необхідних для синтезу РНК.